
Dynamic changes in fruit growth and development of Actinidia ‘Jinyuan’ in two regions
-
摘要:
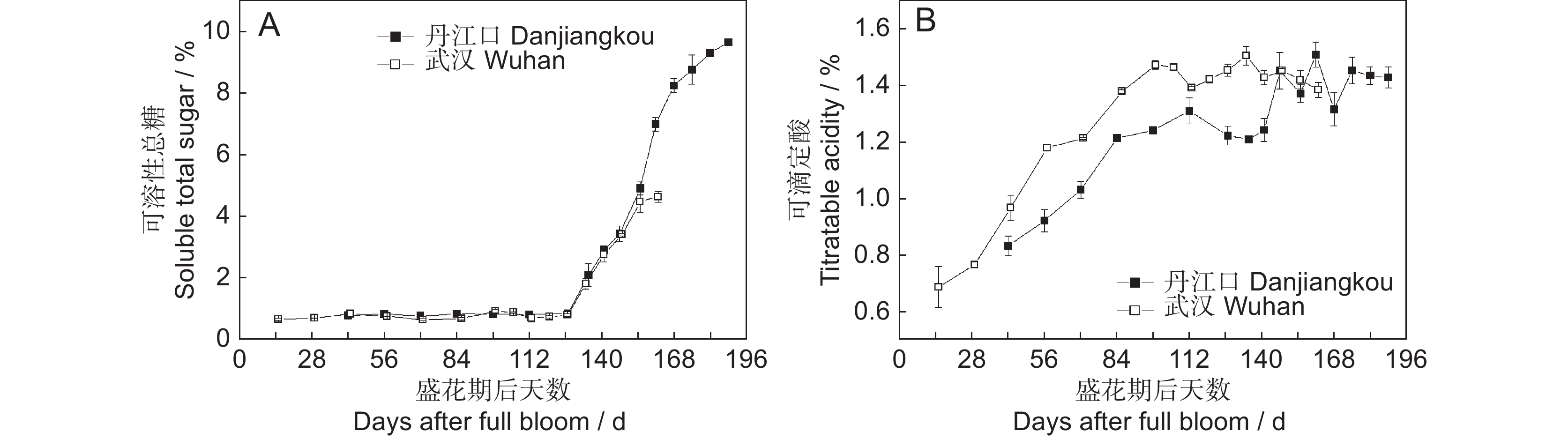
以丹江口和武汉两个地区生产的猕猴桃(Actinidia)品种‘金圆’为研究对象,对果实单果重、纵横侧径、干物质、可溶性固形物、硬度、果肉颜色、可溶性总糖和总酸等指标进行了测定与分析。结果显示:两地果实的纵横侧径和单果重均表现出快速增长-缓慢增长-增长停滞的变化趋势,其中盛花期后57 d内为快速增长阶段。干物质、可溶性固形物、果肉颜色和硬度等果实成熟度指标在两个地区生长期内均表现出高度相似的变化趋势,但是武汉产的果实干物质含量略低于丹江口果实,其硬度又高于丹江口果实。干物质至盛花期后135 d时上升至初步稳定水平;可溶性固形物和可溶性总糖均在盛花期后127 d才开始快速上升,至盛花期后148 d时分别达到约8%和3.5%;果实硬度和果肉颜色均表现为逐渐下降的趋势,其中果实硬度至盛花期后168 d开始急剧下降,果肉颜色至盛花期后155 d下降至103 °hue。综合考虑,‘金圆’果实的适宜采收期应该从盛花期后150 ~ 155 d开始。
Abstract:To elucidate the growth and development of the Actinidia cultivar ‘Jinyuan’ and thus provide guidelines for the development of supported cultivation technology and determination of proper harvest time, fruit grown in the Danjiangkou (DJK) and Wuhan (WH) orchards were regularly measured and analyzed in regard to fruit weight, size, dry matter, soluble solids concentration (SSC), firmness, flesh color, soluble sugar, and total acidity. Results indicated that the size (vertical, horizontal, and lateral diameter) and weight of the fruit grown in both orchards showed a rapid-slow-cease growth trend, and the rapid growth stage mainly occurred within 57 d after full bloom (DAFB). Maturity indices, including dry matter, SSC, flesh color, and firmness, revealed strongly similar patterns during the whole growth period in both orchards, although dry matter was slightly lower and firmness was slightly higher in fruit from WH than from DJK. Dry matter increased to a primary plateau at 135 DAFB. Both SSC and soluble sugar began to increase rapidly from 127 DAFB and reached 8% and 3.5%, respectively, at 148 DAFB. Both fruit firmness and flesh color gradually declined with growth time, with firmness beginning to decline largely at 168 DAFB and flesh color decreasing to 103° hue at 155 DAFB. Taken together, ‘Jinyuan’ fruit should be properly harvested at 150–155 DAFB.
-
Keywords:
- Actinidia /
- ‘Jinyuan’ /
- Fruit /
- Growth and development /
- Maturity
-
山地地区既是生物多样性和遗传多样性的宝库,也是维持全球陆地生态系统平衡重要的组成部分[1, 2]。当今山地生物多样性的形成与演化受长时间尺度地质运动、气候变迁、地貌变化和生态过程的影响,最终落脚于物种形成、适应性进化、拓殖、物种存续和灭绝事件的综合过程[2]。值得注意的是,在大部分生物多样性富集的山地生态系统中都具有明显的垂直地带性差异,即从低海拔分布的森林生物区到高海拔分布的高寒生物区。其中,高寒生物区的形成与山体隆升的幅度及历史环境的变迁密切相关,对认识山地生物多样性形成具有关键性的意义。
高寒生物区位于天然树线以上,永久雪线以下,在各大洲的高山中都有分布,面积约占全球陆地面积的3%,其海拔分布范围因所处纬度不同,自低纬度向高纬度海拔逐渐降低,与各地区树线以及山峰冰川和雪线的高低密切相关(图1)。就植被类型而言,高寒生物区主要包括高山灌丛,草甸和冰缘带植被等[3]。由于低温(昼夜温差大)、大风、空气稀薄、紫外辐射强烈等独特的环境因素影响,高山植物普遍表现出与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性,其植物区系组成主要是适应冰雪或严寒生境的低矮或匍匐状的灌木、禾草型植物、多年生草本、莲座状植物和各种类型的垫状植物[3]。据估计,全球高寒生物区大概有8 000 ~ 10 000种高等植物,分属于大约100个科和2 000余个属,占已知高等植物总数的4%左右[3]。虽然就植物多样性而言,高寒生物区植物种类比热带、亚热带雨林低,但因其自然环境严酷,植物区系极其特殊,同时对全球变暖非常敏感,生态系统极其脆弱,全球气候的变暖将使树线升高,压缩该植被带的范围,使物种灭绝的风险大大增加[4, 5],因此长期以来都是生物和生态学家关注的焦点。
![]() 图 1 高寒生物区分布图A:高寒生物区在全球的分布(平均海拔3 000 m左右),苔原用蓝色表示。全球SRTM 500 m DEM数据和60ºN以上的高程数据来自30弧秒分辨率全球多分辨率地形高程数据2010(GMTED2010);苔原依据WWF的划分标准。B:从赤道到两极主要山脉的高寒生物区海拔高度变化示意图(修改自Körner [3])。Figure 1. Distribution map of world alpine biomeA: Alpine regions are shown in color, tundra regions are shown in blue and non-alpine regions are shown in different degrees of gray. SRTM 500 m and latitude above 60°N are derived from the Global Multi-resolution Terrain Elevation Data 2010 (GMTED2010) at 30 arc-second resolution. Tundra is depicted based on WWF global biome classification. B: Schematic of altitudinal position of alpine life zone from Arctic to Antarctic latitudes (modified from Körner[3]).
图 1 高寒生物区分布图A:高寒生物区在全球的分布(平均海拔3 000 m左右),苔原用蓝色表示。全球SRTM 500 m DEM数据和60ºN以上的高程数据来自30弧秒分辨率全球多分辨率地形高程数据2010(GMTED2010);苔原依据WWF的划分标准。B:从赤道到两极主要山脉的高寒生物区海拔高度变化示意图(修改自Körner [3])。Figure 1. Distribution map of world alpine biomeA: Alpine regions are shown in color, tundra regions are shown in blue and non-alpine regions are shown in different degrees of gray. SRTM 500 m and latitude above 60°N are derived from the Global Multi-resolution Terrain Elevation Data 2010 (GMTED2010) at 30 arc-second resolution. Tundra is depicted based on WWF global biome classification. B: Schematic of altitudinal position of alpine life zone from Arctic to Antarctic latitudes (modified from Körner[3]).高寒生物区作为山地地区海拔最高的生态系统,其形成与区域造山运动和气候变化密切相关,尤其在中低纬度地带,只有山体隆升到一定高度才有可能形成高寒生境,为高寒植物的拓殖和分化创造条件。例如,安第斯Páramos起源时间大概在3.5 Ma左右,与安第斯山北部地区隆升至现在高度的时间一致,同时受到第四纪冰期的塑造作用[6];新西兰的南阿尔卑斯山地区的高寒生物区不早于5 Ma[7, 8],研究表明能够适应高寒生境的类群形成始于山体抬升到一定高度并且气候变冷时,到1.9 Ma 高寒生境广泛且持续存在时,新西兰高寒生物才开始迅速多样化[7, 8];而高寒植物拓殖到东非高寒生物区的时间大概在5-10 Ma,这个时间晚于许多东非高山在渐新世的造山运动,其现今高寒植物多样性格局的形成主要受到更新世气候波动的影响[9]。泛青藏高原地区是全球温带高寒植物多样性最高的区域,也是山地生物多样性研究的热点和难点区域。长期以来,该地区的生物学研究受到早期地质学上整体抬升或近期快速隆升观点的影响,忽视了地域的差异性以及对最新地质学观点的吸纳,随着对青藏高原各地块构造演化历史研究的加深,我们对该地区生物多样性形成与演化的认识也有了长足的进步。青藏高原在地质历史时期经历了复杂而大规模的构造环境变化[10, 11],这一系列的地质运动不仅塑造了该地区复杂的地形地貌,引发了东亚季风系统的改变[12-14],也深刻影响着该地区植物多样性的形成与发展。目前,该地区的生物多样性研究已经逐步迈入到地质-气候-生物过程的交叉研究阶段[15, 16],未来多学科的贯穿融合也必将成为深入揭示该地区生物多样性演化过程及其形成机制的重要手段。
基于此,本文在综合考虑诸如造山运动和气候变化等地球物理过程如何影响高寒物种的积累和多样化过程,以及自然选择和杂交等在维持其生物多样性中发挥的作用,概述了青藏高原及其周边地区高寒植物多样性的起源和演化节奏、成分来源和区系交流及其内外驱动机制等方面的重要研究进展。
1. 青藏-喜马拉雅-横断山地区环境异质性和高寒植物多样性
1.1 青藏-喜马拉雅-横断山地区地质历史背景及现代地理环境差异
从地质构造上看,泛青藏高原并非整体抬升,它由来源和抬升历史不同的地块组成(图2)[17, 18]。构成青藏高原核心的羌塘地块和拉萨地块均来自南半球的冈瓦纳大陆,冈瓦纳大陆裂解后这两个地块北漂,分别于晚三叠世和早白垩世拼合成为欧亚大陆的一部分[19]。随着古高程研究的进一步深入,青藏高原隆升过程的复杂性和块体间差异被逐渐认识。藏南拉萨地块冈底斯山至少在51 Ma 以来就来一直保持着4 500 m以上的海拔[20]。从始新世到晚渐新世,印度-亚洲板块加速汇聚,由板块汇聚导致的地壳挤压增厚使西藏中部和南部进一步抬升[21]。直到新近纪,青藏高原主体中部高地才完全形成[22],到了中中新世,喜马拉雅才隆升至接近现在的高度[23]。横断山一直被认为是在晚中新世到上新世形成的[24],然而,最新的化石和构造证据表明,横断山部分地区在早渐新世就已经达到了接近现代的海拔高度[25]。考虑到高原腹地、横断山和喜马拉雅在形成时间、隆升节奏、地形地貌和气候条件等差异对区域生物多样性的形成与演化的影响[18],有必要将高原腹地、喜马拉雅和横断山区这3个地区区分来看[18, 26, 27](图2)。
![]() 图 2 泛青藏高原相关地质构造及生物地理单元的划分—青藏高原腹地、喜马拉雅和横断山Figure 2. Geological units related to orogeny of Pan-Tibetan Highlands, divided into Tibetan Plateau, Himalaya, and Hengduan Mountains
图 2 泛青藏高原相关地质构造及生物地理单元的划分—青藏高原腹地、喜马拉雅和横断山Figure 2. Geological units related to orogeny of Pan-Tibetan Highlands, divided into Tibetan Plateau, Himalaya, and Hengduan Mountains青藏高原腹地的范围是指由广大高原面构成的主体,是阿尔金断裂以南,雅鲁藏布江缝合带以北,喀喇昆仑山脉走滑断层以东,秦岭、横断山以西,包括昆仑-祁连盆岭(山-盆)区[18, 26, 28]。高原腹地深处大陆内部,且高山环绕,在漫长的地质历史中经历了从湿润-半湿润-半干旱-干旱-极端干旱的气候转变[29],演化成今天包含多种生活型的高寒生态系统[30, 31],一般从东南向西北地势越高种类越少,大体上以草甸灌丛、荒漠草原和冰缘带植被为主,局部地区仍有小面积的森林和零星的森林树种分布[30]。
横断山地处广义青藏高原的东南缘,地域涵盖滇西北、川西、藏东和藏东南延伸至青海东南部和甘肃西南部,由数列南北近平行的山脉组成[26, 32],区域内河流深切,山体平均海拔从川西藏东的4 000 ~ 5 000 m到云南南部地区的2 000 m左右,其中5 000 m以上的极高山和3 500 ~ 5 000 m的高山占到总面积的73%,最高峰贡嘎山的海拔为7 556 m,而山脚海拔仅为1 100 m,岭谷之间相对高差悬殊[33],是世界上特殊环境类型最多的地域之一[34]。与寒旱少雨的青藏高原腹地相比[29],东南部的横断山地区受西南季风及东南季风的双重影响夏季多雨湿润,且高山峡谷逶迤南北,悬殊的谷、岭高差,导致其气候、植被和土壤巨大的垂直分异[35, 36]。由于其地理位置特殊及生境复杂多样,其南端与东南亚热带北缘相邻接,北部及西北部向青藏高原高寒地带过渡,各类植物区系成分交错混杂,形成了齐全的从河谷至高山的垂直植被带。
喜马拉雅山位于青藏高原南部,由数条大致东西向平行的支脉组成并向南凸出呈弧形。它西起克什米尔的南迦-帕尔巴特峰,东至雅鲁藏布江大拐弯处的南迦巴瓦峰,是印度河-雅鲁藏布江缝合带(也是印度板块和欧亚板块的界限)以南,主边界断层以北的地区(图2)[26, 27]。其南北向的断裂构造发育,经河流切割形成纵向深险峡谷,成为西南季风气流北进的通道。喜马拉雅山西部紧挨着兴都库什山系,东边紧靠印度-缅甸,北部紧靠喀喇昆仑山脉和冈底斯山。由于其独特的地理位置及周围毗邻地区生态环境的显著差异,喜马拉雅植物区系包含了许多不同的地理成分(东亚成分,马来-缅甸成分和中亚细亚及地中海成分等),植物区系和植被景观也呈现出明显的垂直地带性,其中热带成分在低海拔地区占优势,东亚温带成分则在亚高山和高山地带占优势,该地区不存在真正的特有科,特有属也不多[37],特有种主要在中高海拔地区[38]。
1.2 青藏-喜马拉雅-横断山地区高寒植物多样性
受区域地质构造历史及其环境异质性的影响,青藏高原腹地、喜马拉雅和横断山的高寒植物物种的丰富度和演化历史也表现出区域间的差异性和交融性。其中,横断山的高寒生物多样性尤其丰富。根据张大才和孙航[39]统计,在横断山4 100 m以上的海拔范围共有种子植物70科297属1 820种,而这一数字很可能低估了横断山高寒地区的植物多样性,因为横断山地区的树线在很多山区可下探至3 500 m左右(图3)。据最近的统计结果,除亚种和变种外,横断山高寒地区种子植物可达3 030种,其中包括12种裸子植物[40],远高于面积更广阔的青藏高原腹地(种子植物1883种)[30, 41],和喜马拉雅地区(据统计,以尼泊尔为代表的中喜马拉雅地区共有高寒种子植物1 227种,而西喜马拉雅则记录有约830种)[42, 43]。在植物区系的研究中,武素功等[30]指出,青藏高原主体的高寒植物区系属于中国-喜马拉雅区系的衍生或组成成分,缺乏古特有属,大部分类群是从分布较广、尤其是横断山和青藏高原低海拔的一些属种衍生而来。
![]()
2. 青藏-喜马拉雅-横断山地区高寒植物多样性演化历史
2.1 青藏-喜马拉雅-横断山地区高寒植物多样性的起源及演化节奏
从全球高寒植物多样性的形成历史来看,高寒植物区系是一个比较年轻的生物区,主要受到晚中新世以来各大洲山体最后阶段的抬升和第四纪冰期的驱动[7, 45, 46]。然而,相对于其他高寒生物区,青藏高原-喜马拉雅-横断山地区的高寒植物多样性更为丰富也更加古老,受到区域构造隆升和气候变化的强烈影响。
近几年,在青藏高原及其周边地区基于关键类群和特有类群在分子水平上开展了大量的谱系地理和生物地理学研究,为认识该地区重要类群的起源和高寒植物多样性演化发挥了重要作用。例如,Luo 等[47]对横断山-喜马拉雅冰缘带特有的4个物种的谱系地理分析结果表明,4种植物主要分化于晚上新世至早-中更新世;一些特有种比例较高的属,例如,毛冠菊属(Nannoglottis)、川木香属(Dolomiaea)、重羽菊属(Diplazoptilon)、黄缨菊属(Xanthopappus)、无心菜属(Arenaria)中的垫状类群以及高山竹类也是在上新世-第四纪快速分化[48-52];而橐吾(Ligularia)-垂头菊属(Cremanthodium)-蟹甲草属(Parasenecio)[53]、风毛菊属(Saussurea)[54]、锦鸡儿属(Caragana)[55]、绿绒蒿(Meconopsis)[56]、大黄属(Rheum)[57]和红景天属(Rhodiola)[58]则是在中中新世至晚中新世发生了快速多样化。然而,更古老的拓殖和分化事件仍然发现于其他广布且多样化的类群当中。Ebersbach等[59] 通过虎耳草属(Saxifraga)的生物地理分析把高寒类群拓殖于青藏高原的时间推进到晚始新世(49-34 Ma),与当时西藏南部已经有超过4 000 m的高山相吻合[20];随后的多样化发生在10-4 Ma认为是受到较晚抬升的横断山构造活动的影响。龙胆属(Gentiana)在始新世(40-34 Ma)起源于青藏高原及其周边地区,除了晚中新世-早上新世的突然加快,自晚始新世以来多样化速率增加缓慢[60]。Zhao 等[61]对喜马拉雅-横断山分布的高山姜类的研究发现,大概在早渐新世( ~ 32 Ma)象牙参属(Roscoea)和距药姜属(Cautleya)就与其姐妹群分化开来,而象牙参属在横断山与喜马拉雅的两个分支大概在渐新世-中新世之交分化[61]。从以上案例研究结果可以看出,青藏高原及其周边山地高寒植物的多样化过程基于不同研究类群的演化尺度,表现出了时空上的差异性,分化时间从古老到年轻,时间跨度大。另外,对广义青藏高原的笼统划分也限制了对不同地区演化历史及其驱动机制的深入理解和认识。为了克服这个问题,Ding 等[27]选取了青藏高原腹地,喜马拉雅和横断山高寒植物多样性较高的18个被子植物类群(总计包含3798种),通过应用系统发育比较方法和多类群大尺度的整合分析,重建了横断山、喜马拉雅以及青藏高原主体高寒植物多样性连续变化的时空演化历史,将横断山高寒植物的起源时间追溯到晚始新世到早渐新世[27],并结合古生物学和地质学证据,探讨了重大地质构造事件(山体隆升)和历史气候变化对高寒植物多样性形成的影响(图4)。研究表明,在横断山地区,高寒物种就地演化速率在早中新世到中中新世(23-15 Ma)和晚中新世(10-7 Ma)加快并一直保持较高的水平,是新近纪全球降温、横断山造山运动与亚洲季风增强共同作用的结果。而在喜马拉雅地区,物种演化速率的加快比横断山晚,主要在中中新世(19 Ma 一直持续到12 Ma),与喜马拉雅的隆升节奏和夏季季风的增强有关[27]。青藏高原地区高寒植物多样性增加紧随喜马拉雅其后(18 Ma),说明现今青藏高原(腹地)的高寒植物多样性很可能是喜马拉雅隆升后发展起来的[27]。因此,横断山及其周边地区高寒生物多样性的形成是自始新世降温以后,在造山运动和季风加强的共同影响下形成的,这些因素一起为生物多样性演化提供了一个生态-进化的舞台,并且一直持续至今。
![]() 图 4 横断山、喜马拉雅和青藏高原腹地高寒地区生物多样性演化速率与气候变化和地质历史之间的关系(修改自Ding 等[27])A:全球气候变化曲线来自深海氧同位素记录[64, 65]。蓝色线段表示亚洲季风演化趋势,由Farnsworth 等[66] 在理想CO2下模拟的青藏高原及其周边地区各地史阶段的年平均降水量表示。B:喜马拉雅(HIM)、青藏高原腹地(TP)和横断山(HDM)从晚始新世至今分3个阶段的地形示意图。红色带数字的圆点表示基于最新构造证据重建古高程的地点。C:横断山、喜马拉雅和青藏高原腹地高寒生物区植物多样性速率随时间的变化。青藏高原主体图中由浅至深的黄色条带代表了古近纪以来青藏高原腹地的干旱化程度。Figure 4. Rates of biotic assembly in relation to climate and geological history in the Hengduan Mountains, Himalaya, ibetan Plateau (modified from Ding et al.[27])A: Evolution of global climate is represented by deep-sea oxygen-isotope records[65] and estimated deep ocean temperatures by Hansen et al. [64]. Monsoon conditions are indicated by modeled mean annual precipitation (m) for each geological stage, represented by blue lines at idealized CO2 (solid blue circle) (modified from Farnsworth et al. [66]). B: Schematic of topography of the Himalaya (HIM), the Tibetan Plateau (TP), and the Hengduan Mountains (HDM) in three phases, from late Eocene to the present. C: Rolling estimates of rates through time in the HDM, HIM, and TP. Light to dark yellow bar in the last panel represents intensity of aridification in the TP since the Paleogene.
图 4 横断山、喜马拉雅和青藏高原腹地高寒地区生物多样性演化速率与气候变化和地质历史之间的关系(修改自Ding 等[27])A:全球气候变化曲线来自深海氧同位素记录[64, 65]。蓝色线段表示亚洲季风演化趋势,由Farnsworth 等[66] 在理想CO2下模拟的青藏高原及其周边地区各地史阶段的年平均降水量表示。B:喜马拉雅(HIM)、青藏高原腹地(TP)和横断山(HDM)从晚始新世至今分3个阶段的地形示意图。红色带数字的圆点表示基于最新构造证据重建古高程的地点。C:横断山、喜马拉雅和青藏高原腹地高寒生物区植物多样性速率随时间的变化。青藏高原主体图中由浅至深的黄色条带代表了古近纪以来青藏高原腹地的干旱化程度。Figure 4. Rates of biotic assembly in relation to climate and geological history in the Hengduan Mountains, Himalaya, ibetan Plateau (modified from Ding et al.[27])A: Evolution of global climate is represented by deep-sea oxygen-isotope records[65] and estimated deep ocean temperatures by Hansen et al. [64]. Monsoon conditions are indicated by modeled mean annual precipitation (m) for each geological stage, represented by blue lines at idealized CO2 (solid blue circle) (modified from Farnsworth et al. [66]). B: Schematic of topography of the Himalaya (HIM), the Tibetan Plateau (TP), and the Hengduan Mountains (HDM) in three phases, from late Eocene to the present. C: Rolling estimates of rates through time in the HDM, HIM, and TP. Light to dark yellow bar in the last panel represents intensity of aridification in the TP since the Paleogene.2.2 青藏-喜马拉雅-横断山地区高寒植物的成分来源和区系交流
横断山和喜马拉雅的高寒植物多样性积累均以就地演化为主,而高原腹地则以迁入为主。此外,横断山高寒生物区和喜马拉雅、高原腹地之间还有着非常密切的区系联系,是喜马拉雅和青藏高原高寒生物多样性的主要供给地[27]。相比之下,横断山中低海拔要远高于其邻近高寒生物区对横断山高寒生物区的贡献,这可能与横断山整体上的温带植物区系性质有关,横断山也被认为是北温带分布型属的起源和分化中心[62]。喜马拉雅地处中纬度地区,下接亚热带气候类型,上接温带气候类型,直到早中新世-中中新世以后才快速隆起,而邻近的横断山和青藏高原地区隆升历史较早,尤其是横断山复杂的地形地貌维持了比较高的高寒生物多样性[62],因此,已经预适应高寒的横断山类群是喜马拉雅高寒生物区的重要来源。Hörandl和Emadzade[63]通过对毛茛属(Ranunculus)的生物地理研究也得出喜马拉雅地区的高寒类群,相对于从其他地方通过长距离扩散迁移过来(例如北极地区,中亚和北亚以及从台湾高山地区),从中低海拔就地演化而来的较少。另外,植物区系调查结果表明,喜马拉雅的特有成分主要集中在高寒/亚高山地带[38]。由此可见,喜马拉雅的快速隆升所形成的高寒地带为随后的迁移和就地演化创造了条件。由于隆升过程始终占主导地位,因而可以像横断山一样的温带成分得到极大地发展。
更新世冰期时,由于山岳冰川发育,树线比现在低,邻近山地高寒生物区之间的交流频率增加[67]。即使是位于赤道附近的埃塞俄比亚高地和乞力马扎罗山在冰期时高寒生物区的海拔也比现在低1 000~1 500 m[68],使得冰期时高寒生物区之间的联系更加密切。在以往的谱系地理研究中也发现,虽然不同物种在冰期、间冰期的扩张方式不同,但都多次出现从横断山回迁到青藏高原腹地的情况[69]。Ding 等[27]研究也发现,贯穿第四纪从横断山扩散到其周边地区,尤其是迁移到青藏高原的速率快速增加。
2.3 青藏-喜马拉雅-横断山地区高寒植物多样化的内在驱动力
长期以来的地质活动(山体隆升、河流重组等)和气候演变不断地塑造着青藏高原及其周边地区的地形地貌,由此产生的新生境、局域环境的改变和生态位的分化为物种形成、演化和迁移创造了机会[27, 70]。较高的生境异质性和气候的反复波动不仅促进了不同居群间的隔离和遗传分化[71],沿海拔梯度不同强度的自然选择压力以及局域微生境的多样化也使得有限的区域内居群承受的生态选择压力不同,进而导致生殖隔离的产生和同域物种形成[72]。大量群体基因组学的分析还发现基因流在青藏高原及其周边山地很多植物类群的物种形成过程中普遍存在,这可能受到第四纪气候震荡或者河流袭夺事件的影响,使得尚未分化完全的物种有了再次接触和基因交换的机会[73]。在一些多样化较高的类群中,如杜鹃花属(Rhododendron)[74]、龙胆属[75]等,频繁发生的种间杂交也为新物种的形成创造了机会,表明很多物种仍处于物种分化的过程当中[73]。同时,越来越多的证据指出异源多倍体对生物多样性的贡献很可能被低估。例如,蔊菜属(Rorippa)中高蔊菜(Rorippa elata (Hook. f. & Thomson) Hand.-Mazz.)和沼生蔊菜(Rorippa palustris (L.) Besser)的杂交起源及它们在横断山区从南到北反复拓殖的群体历史,表明多倍体物种种群扩张过程中种群的适应能力呈现出沿纬度梯度提高的趋势,多倍体可以通过调整适应性来应对扩张过程中的选择压力[76]。因此,生物因素(杂交,多倍化,基因渐渗)和生态过程(自然选择,适应性进化)作为物种形成和分化的内在驱动力,与构造运动和气候变化等外营力共同作用,在维持该区域遗传多样性和生物多样性上扮演着重要角色。
3. 展望
随着基于近几年对泛青藏高原构造演化历史及其所引发的气候演变的认识逐步加深,本文以不同地理单元的划分作为切入点,归纳并总结了横断山、喜马拉雅和高原腹地高寒植物多样性演化历史、过程及其形成机制的重要研究进展。首先,根据各地的环境差异和构造背景界定不同的生物地理区,如,横断山、喜马拉雅和青藏高原腹地的划分,进而比较其多样性形成历史的异同,在一定程度上解决了长期以来因为地域界定问题由不同类群推演出来生物地理历史和多样性成因互存争议的局面。此外,本文还阐述了杂交、多倍化和基因渐渗等生物过程在活跃的构造运动和气候波动下维持区域生物多样性的重要作用。
青藏高原及其周边山脉活跃的构造抬升、季风气候的加强以及更新世的冰川作用等环境变化深刻塑造了泛青藏高原的地形地貌和气候环境的异质性,为该地区的物种分化和形成创造了条件。自然选择、适应性进化和杂交作为物种多样性形成的内在驱动力,在维持该区域遗传多样性和生物多样性上也扮演着重要角色。泛青藏高原地区独特的地理环境和复杂的构造演化背景,在一定程度上对我们深入揭示其多样性的形成和维持机制提出了挑战,也正因如此,未来该地区的生物多样性研究势必推动生物学和地质学等多学科的交叉融合,通过地质-气候-生物的过程的耦合关系和相互作用的研究加深对地球环境变化和生物演化过程的认识。
不同山地地区现代环境、地形地貌差异显著,地质背景和环境演化历史迥异,其生物多样性的演化历史必然也呈现出时空格局上的差异。深入比较不同山地生物多样性形成演化历史的异同及与各自环境变化的关系、揭示其间的联系无疑将加深对现今山地生物多样性分布格局的理解。例如,以欧亚山地生态系统为代表的横断山、喜马拉雅、天山、高加索山和阿尔卑斯山,它们彼此在共有植物科属组成上都具有一定相似性,包括了以菊科、禾本科、十字花科、豆科、毛茛科、石竹科、蔷薇科、莎草科、虎耳草科、景天科、报春花科、紫堇科、龙胆科、百合科以及一些灌木类群,如杜鹃花科、小檗科、杨柳科等为代表的一些高寒植物类群,组成了欧亚大陆甚至北半球高寒生物区最主要的植物种类和多样性[3, 31, 77]。不同区域的构造演化和气候变化历史是否影响了它们之间的物种交流,又是否导致了这些山地地区间生物多样性的形成、演化历史的差异?利用这些在不同山区广泛分布的特征植物类群,比较其物种多样化的过程和联系,将有助于回答以上问题。此外,高寒生物区严苛的自然环境所造成的选择压力是否导致了不同类群在功能形态上的趋同演化或繁殖策略的改变,例如,与高寒植物的适应性进化策略相关的某些关键形态性状—温室结构、棉毛结构及垫状结构,这些特殊的生活型或生理代谢等有时会同时出现在亲缘关系较远的类群中。把这些关键性状的演化与特定生物区的演化结合起来分析并探究其适应性和内在遗传机制,将会进一步加深我们对生物多样性在不同时间尺度和驱动因素下演变的认识。
-
![]()
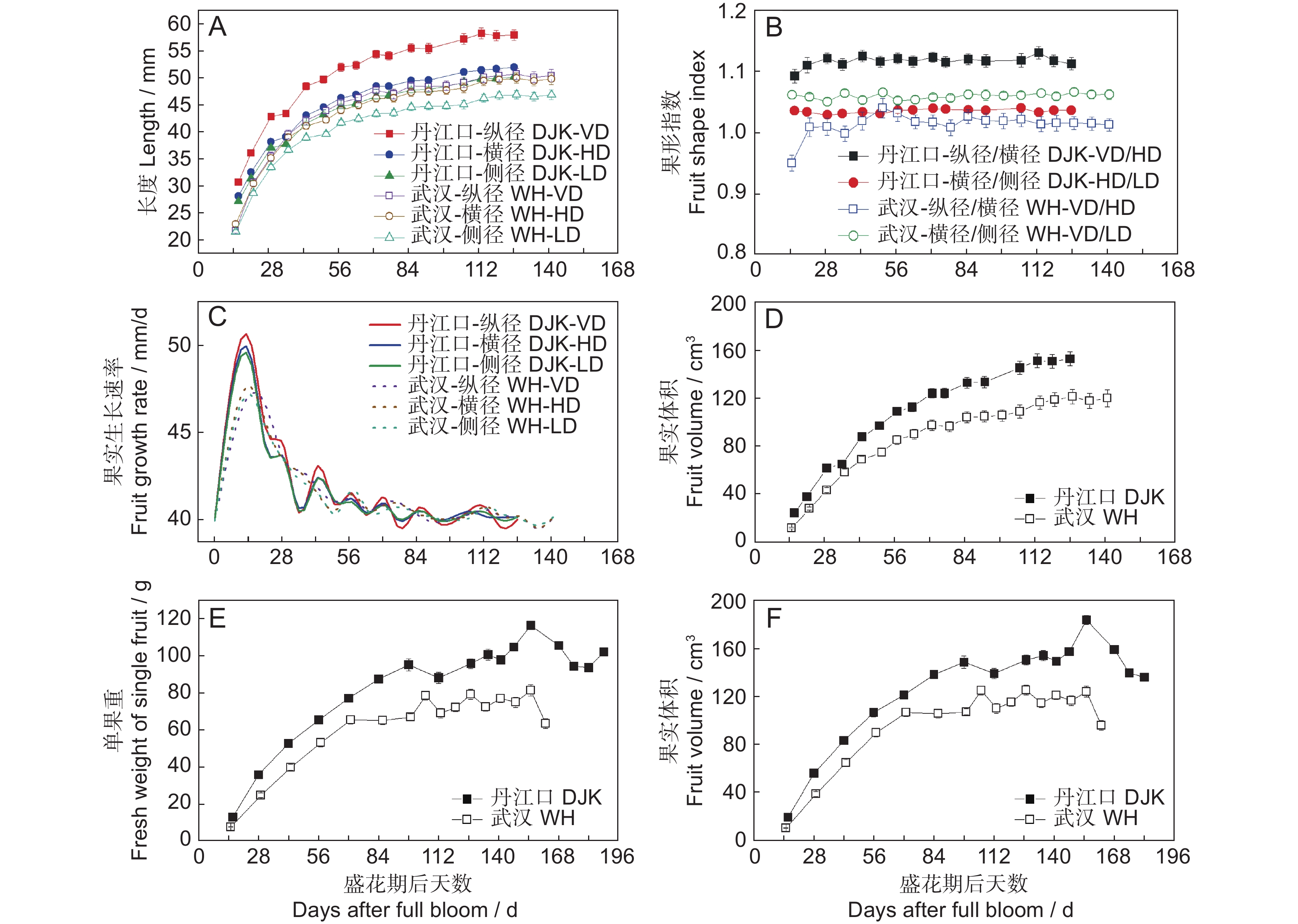
图 1 丹江口和武汉生长的‘金圆’果实生长趋势
A ~ D为树上固定果实的生长趋势。A:果实纵横侧径;B:果形指数;C:纵横侧径的生长速率;D:果实体积;E、F:下树后果实的生长趋势(E为单果重,F为果实体积)。
Figure 1. Growth trends of ‘Jinyuan’ fruit on and off vine grown in Danjiangkou (DJK) and Wuhan (WH) orchards
A–D indicate growth of fruit fixed on vine during whole period. A: Length of fruit vertical diameter (VD), horizontal diameter (HD), and lateral diameter (LD); B: Fruit shape index based on ratio of VD/HD and HD/LD; C: Growth rates of VD, HD, and LD; D: Fruit size based on VD, HD, and LD data. E and F indicate growth of fruit detached from vine during monitoring period (E, fresh weight of single fruit; F, fruit volume calculated from VD, HD, and LD data).
![]()
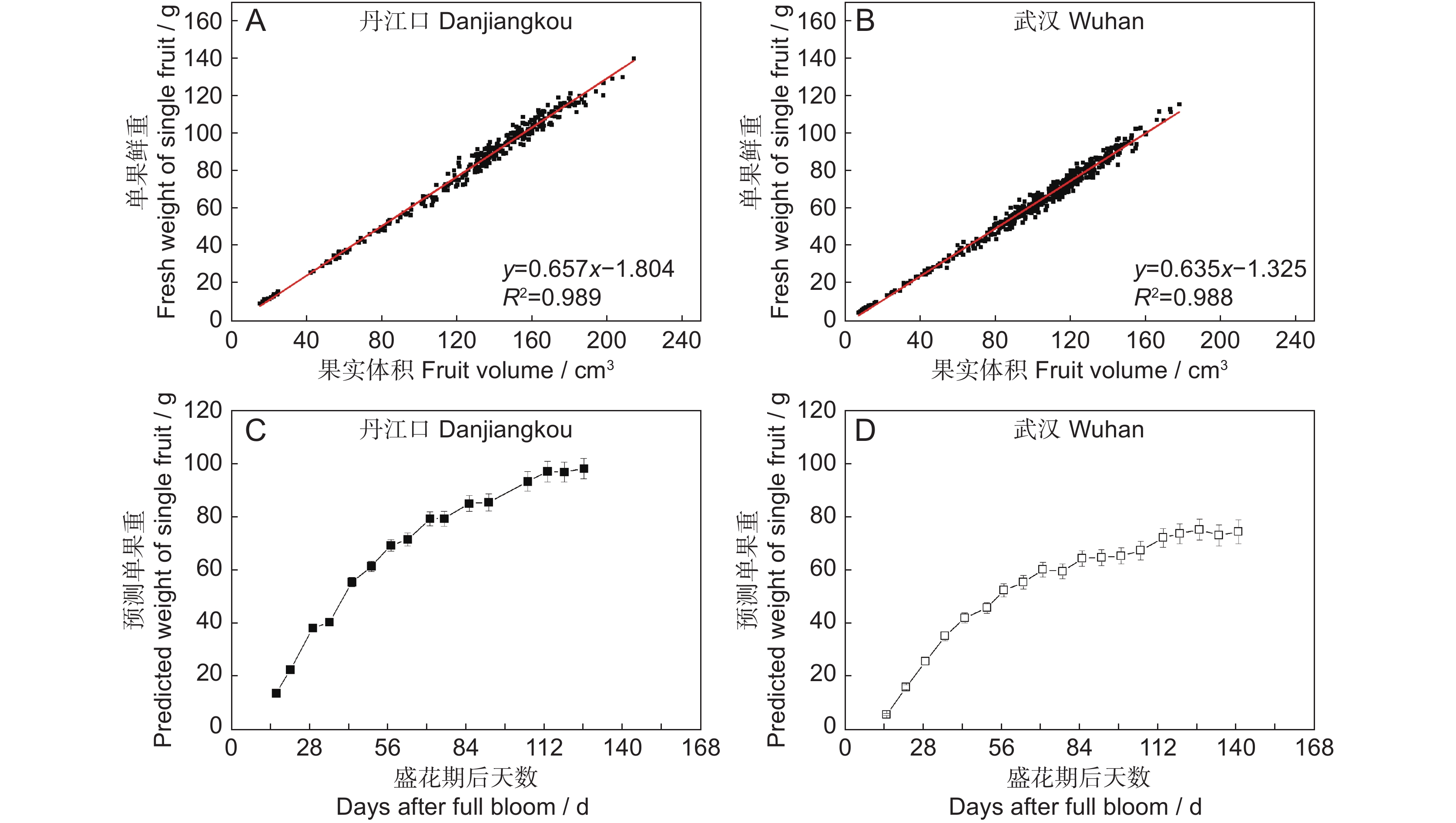
图 2 ‘金圆’果实重量与果实体积的相关性分析及预测单果重变化
A、B:丹江口(A)和武汉(B)生产的‘金圆’果实重量与体积的相关性分析; C、D:丹江口(C)和武汉(D)生产的‘金圆’在生长期内的预测单果重变化。单果重的预测基于图1中固定在树上的果实体积数据。
Figure 2. Correlation analysis of fruit weight and volume and change in predicted weight of ‘Jinyuan’ fruit
A, B: Correlation analysis of fruit weight and volume of ‘Jinyuan’ produced in Danjiangkou (A) and Wuhan orchards (B); C, D: Change in predicted weight of ‘Jinyuan’ fruit on vines during growth period in Danjiangkou (C) and Wuhan (D) orchards. Volume data of fruit fixed on vines in Fig. 1 were used to predict fruit weight.
![]()
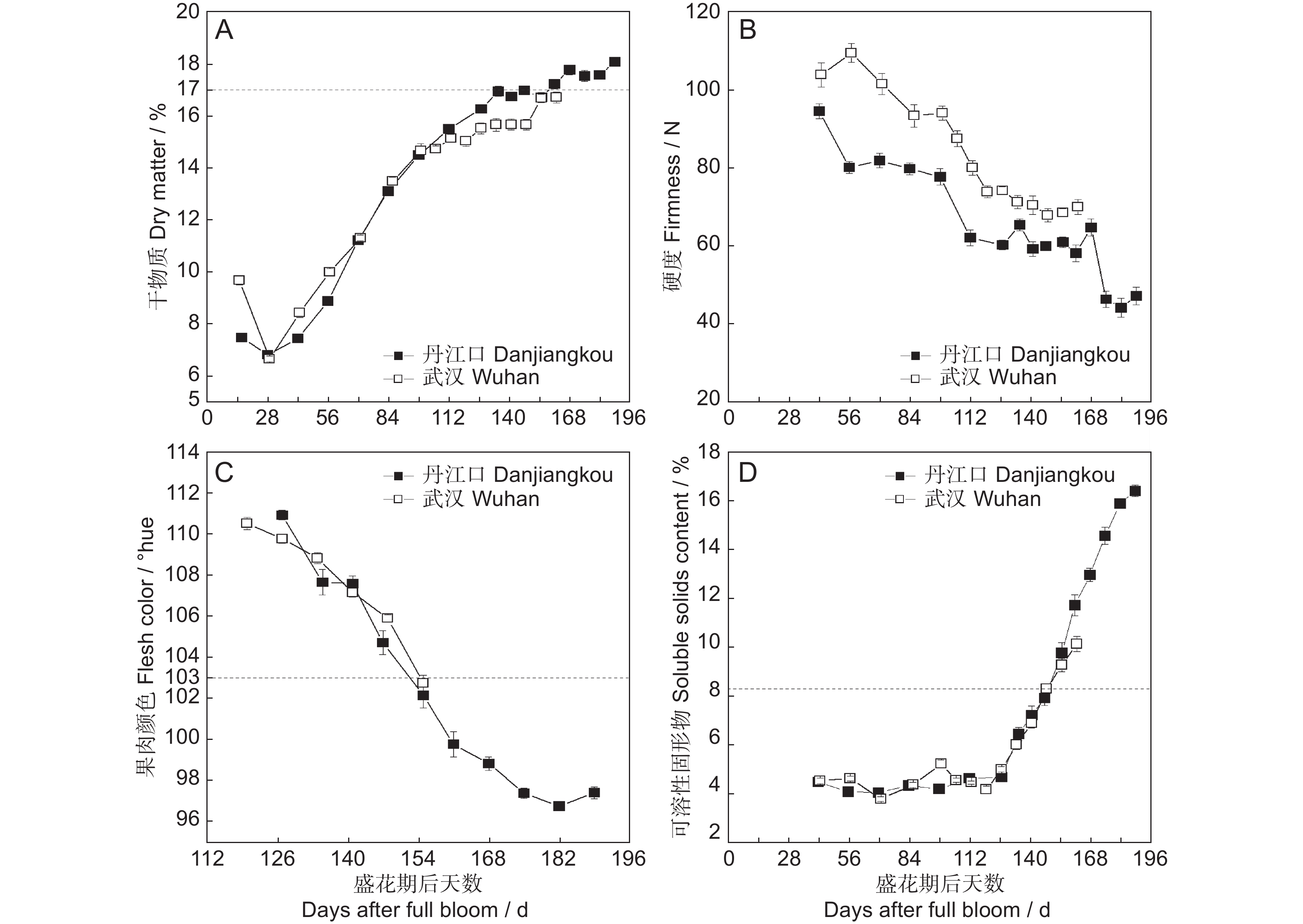
图 3 丹江口和武汉产的‘金圆’果实成熟度变化
A:干物质;B:硬度;C:果肉颜色;D:可溶性固形物。
Figure 3. Maturity changes in ‘Jinyuan’ fruit produced from Danjiangkou and Wuhan orchards during growth
A: Dry matter; B: Firmness; C: Flesh color; D: Soluble solids content.
![]()
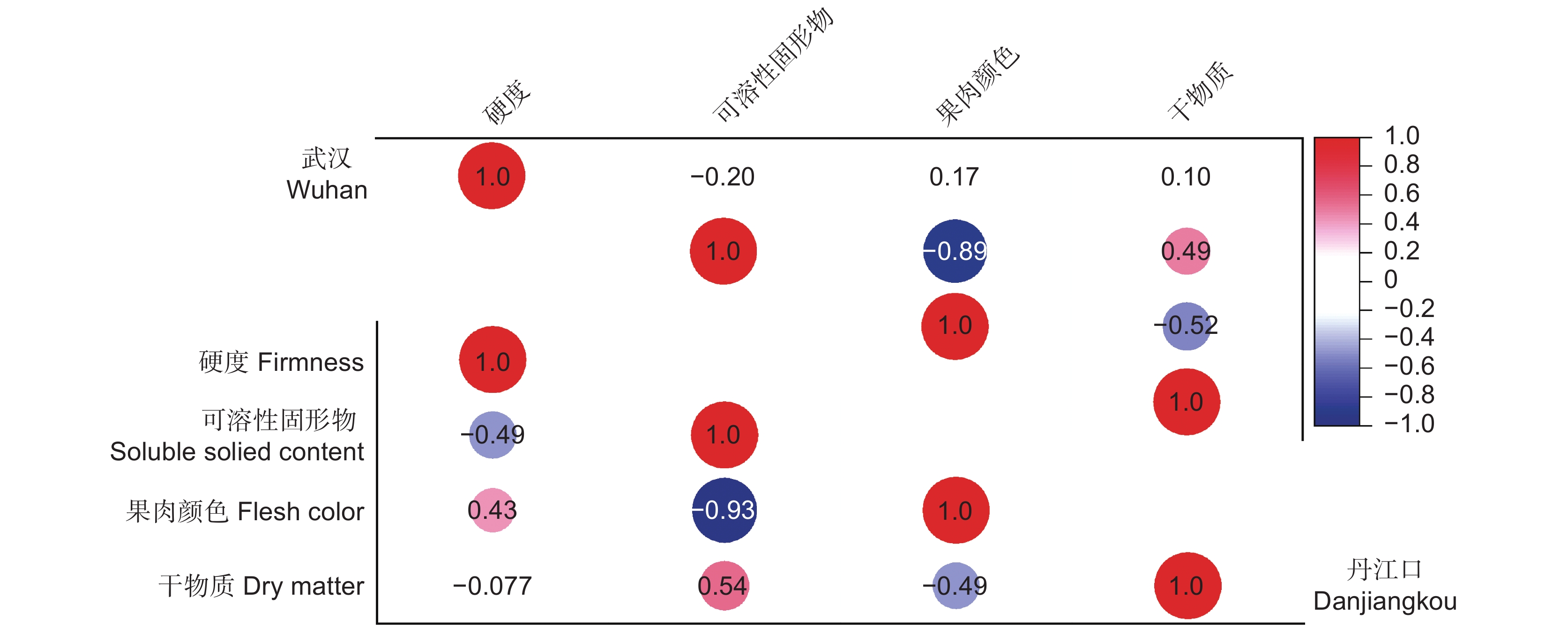
图 4 丹江口和武汉产的‘金圆’果实成熟度指标之间的相关性系数
供相关性分析的各成熟度指标数据来源于果实近成熟期内的数据,丹江口为盛花期后127 ~ 189 d内(n = 209),武汉为盛花期后120 ~ 162 d内(n = 140)。
Figure 4. Correlation coefficients among maturity indices of ‘Jinyuan’ fruit grown in Danjiangkou and Wuhan orchards
Data used for correlation analysis were derived from the near-maturity period of fruit growth, 127–189 DAFB for Danjiangkou fruit (n = 209) and 120–162 DAFB for Wuhan fruit (n = 140).
-
[1] 黄宏文. 猕猴桃属分类资源驯化栽培[M]. 北京: 科学出版社, 2013: 2. [2] Zhong CH,Huang WJ,Wang ZP,Li L,Li D,et al. The breeding progress and development status of the kiwifruit industry in China[J]. Acta Hortic,2022,1332:445−454.
[3] Zhong CH,Wang SM,Jiang ZW,Huang HW. ‘Jinyan’,an interspecific hybrid kiwifruit with brilliant yellow flesh and good storage quality[J]. HortScience,2012,47 (8):1187−1190. doi: 10.21273/HORTSCI.47.8.1187
[4] 韩飞,黄宏文,李大卫,张琼,姜正旺,等. 优质鲜食黄肉猕猴桃新品种‘金圆’的选育及特性分析[J]. 植物科学学报,2019,37(2):171−180. Han F,Huang HW,Li DW,Zhang Q,Jiang ZW,et al. Breeding and characterization of a new high-quality yellow-fleshed kiwifruit cultivar ‘Jinyuan’[J]. Plant Science Journal,2019,37 (2):171−180.
[5] Salinero MC,Vela P,Sainz MJ. Phenological growth stages of kiwifruit (Actinidia deliciosa ‘Hayward’)[J]. Sci Hortic,2009,121 (1):27−31. doi: 10.1016/j.scienta.2009.01.013
[6] Hall AJ,Richardson AC,Snelgar WP. Modelling fruit development in ‘Hayward’ kiwifruit[J]. Acta Hortic,2006,707:41−48.
[7] 金方伦,韩成敏,黎明. 贵长猕猴桃果实生长发育规律研究[J]. 贵州农业科学,2010,38(5):180−183. doi: 10.3969/j.issn.1001-3601.2010.05.052 Jin FL,Han CM,Li M. Fruit growth and development regulation of Guichang gooseberry[J]. Guizhou Agricultural Sciences,2010,38 (5):180−183. doi: 10.3969/j.issn.1001-3601.2010.05.052
[8] 杨朋燕,姚春潮,李小莹,董慧,乔金梅. “徐香”猕猴桃果实生长发育规律的研究[J]. 北方园艺,2014(18):47−50. Yang PY,Yao CC,Li XY,Dong H,Qiao JM. Study on the growth and development regularity of ‘Xuxiang’ kiwifruit[J]. Northern Horticulture,2014 (18):47−50.
[9] Richardson AC,Boldingh HL,McAtee PA,Gunaseelan K,Luo ZW,et al. Fruit development of the diploid kiwifruit,Actinidia chinensis ‘Hort16A’[J]. BMC Plant Biol,2011,11:182. doi: 10.1186/1471-2229-11-182
[10] 王琪凯,杨丹,张晓琴,张信旺. ‘金艳’猕猴桃果实生长动态规律和贮藏性能[J]. 食品科学,2016,37(9):129−133. Wang QK,Yang D,Zhang XQ,Zhang XW. Growth pattern and storage performance of ‘Jinyan’ kiwifruits[J]. Food Science,2016,37 (9):129−133.
[11] 钟彩虹,张鹏,韩飞,李大卫. 猕猴桃种间杂交新品种‘金艳’的果实发育特征[J]. 果树学报,2015,32(6):1152−1160. doi: 10.13925/j.cnki.gsxb.20140494 Zhong CH,Zhang P,Han F,Li DW. Studies on characterization of fruit development of interspecific hybrid cultivar-‘Jinyan’[J]. Journal of Fruit Science,2015,32 (6):1152−1160. doi: 10.13925/j.cnki.gsxb.20140494
[12] 徐明, 雷靖, 刘佩, 李永武, 索江涛, 雷玉山. 中华猕猴桃‘璞玉’果实生长发育动态分析[J/OL]. 分子植物育种, 2022. [2023-01-16]. http://kns.cnki.net/kcms/detail/46.1068.s.20211230.1423.010.html. Xu M, Lei J, Liu P, Li YW, Suo JT, Lei YS. Dynamic analysis of fruit growth and development of ‘Puyu’ kiwifruit[J/OL]. Molecular Plant Breeding, 2022. [2023-01-16]. http://kns.cnki.net/kcms/detail/46.1068.s.20211230.1423.010.html.
[13] 张茜,李明章,王丽华,刘鑫,樊高敏. 黄肉猕猴桃新品种“金实1号”果实生长规律研究[J]. 资源开发与市场,2019,35(6):845−848. Zhang Q,Li MZ,Wang LH,Liu X,Fan GM. Research on fruit growth rule of new yellow flesh kiwifruit cultivar “Jinshi 1”[J]. Resource Development & Market,2019,35 (6):845−848.
[14] 叶发荣,李秀亚,肖钧,宋福兵,莫庆忠,周玉萍. 水城红阳猕猴桃果实生长发育规律[J]. 贵州农业科学,2017,45(11):89−92. doi: 10.3969/j.issn.1001-3601.2017.11.024 Ye FR,Li XY,Xiao J,Song FB,Mo QZ,Zhou YP. Growth and development regularity of Hongyang kiwifruit in Shuicheng[J]. Guizhou Agricultural Sciences,2017,45 (11):89−92. doi: 10.3969/j.issn.1001-3601.2017.11.024
[15] 丁捷,刘书香,宋会会,王瑞玲,秦文. 红阳猕猴桃果实生长发育规律[J]. 食品科学,2010,31(20):473−476. Ding J,Liu SX,Song HH,Wang RL,Qin W. Growth and development patterns of Hongyang kiwifruits[J]. Food Science,2010,31 (20):473−476.
[16] 张茜,李明章,王丽华,刘鑫,樊高敏. 红肉猕猴桃新品种红实2号果实生长规律研究[J]. 湖北农业科学,2020,59(10):97−100. doi: 10.14088/j.cnki.issn0439-8114.2020.10.022 Zhang Q,Li MZ,Wang LH,Liu X,Fan GM. Study on the growth law of the new red flesh kiwifruit cultivar Hongshi No. 2 fruit[J]. Hubei Agricultural Sciences,2020,59 (10):97−100. doi: 10.14088/j.cnki.issn0439-8114.2020.10.022
[17] 韩飞,陈美艳,李昆同,黄文俊,闫春林,等. 不同产地‘金圆’猕猴桃低温贮藏下的生理指标及贮藏性变化[J]. 植物科学学报,2018,36(3):381−392. Han F,Chen MY,Li KT,Huang WJ,Yan CL,et al. Changes in physiological indices and storage properties of ‘Jinyuan’ kiwifruit from different orchards under low temperature storage[J]. Plant Science Journal,2018,36 (3):381−392.
[18] Yuan H,Martin P,Fullerton C,Gould N,Hall A,Burdon J. Refining the estimation of kiwifruit size from linear fruit dimensions[J]. Sci Hortic,2020,262:108878. doi: 10.1016/j.scienta.2019.108878
[19] Li HJ,Pidakala P,Billing D,Burdon J. Kiwifruit firmness:measurement by penetrometer and non-destructive devices[J]. Postharvest Biol Technol,2016,120:127−137. doi: 10.1016/j.postharvbio.2016.06.007
[20] 李秀亚,肖均,许金宽,宋福兵,谭晶晶. 贵长猕猴桃果实生长发育规律[J]. 安徽农业科学,2018,46(21):75−77. doi: 10.3969/j.issn.0517-6611.2018.21.021 Li XY,Xiao J,Xu JK,Song FB,Tan JJ. Growth and development regularity of Guichang kiwifruit[J]. Journal of Anhui Agricultural Sciences,2018,46 (21):75−77. doi: 10.3969/j.issn.0517-6611.2018.21.021
[21] Minchin PEH,Richardson AC,Patterson KJ,Martin PJ. Prediction of final weight for Actinidia chinensis ‘Hort1 6A’ fruit[J]. New Zeal J Crop Hort Sci,2003,31 (2):147−157. doi: 10.1080/01140671.2003.9514247
[22] Burdon J,Pidakala P,Martin P,McAtee PA,Boldingh HL,et al. Postharvest performance of the yellow-fleshed ‘Hort16A’ kiwifruit in relation to fruit maturation[J]. Postharvest Biol Technol,2014,92:98−106. doi: 10.1016/j.postharvbio.2014.01.004
[23] Burdon J,McLeod D,Lallu N,Gamble J,Petley M,Gunson A. Consumer evaluation of “Hayward” kiwifruit of different at-harvest dry matter contents[J]. Postharvest Biol Technol,2004,34 (3):245−255. doi: 10.1016/j.postharvbio.2004.04.009
[24] Crisosto CH,Zegbe J,Hasey J,Crisosto GM. Is dry matter a reliable quality index for ‘Hayward’ kiwifruit?[J]. Acta Hortic,2011,913:531−534.
[25] Burdon JN. Kiwifruit biology: the commercial implications of fruit maturation[M]//Warrington I, ed. Horticultural Reviews: Vol 46. New Jersey: John Wiley & Sons, 2019: 385-421.
-
期刊类型引用(0)
其他类型引用(3)

 下载:
下载:



计量
- 文章访问数: 158
- HTML全文浏览量: 28
- PDF下载量: 31
- 被引次数: 3
