
Coupling relationship between nutrient resorption and stoichiometric ratios of N, P, and K in Setaria viridis (L.) Beauv. leaves
-
摘要:
为探究狗尾草(Setaria viridis (L.) Beauv.)对砂质海岸恶劣环境的适应性,本文以烟台沿海防护林自然保护区核心区内黑松防护林下的狗尾草为研究对象,测定狗尾草成熟叶和衰老叶碳(C)、氮(N)、磷(P)、钾(K)元素的含量,分析其生态化学计量特征和养分重吸收特征,进而研究两者之间的偶联关系。结果显示,狗尾草叶片N∶P > 16,狗尾草生长发育更多受到P元素限制。叶片氮重吸收率(NRE)、磷重吸收率(PRE)、钾重吸收率(KRE)分别为(58.038 ± 2.973)%、(62.167 ± 5.301)%、(86.557 ± 0.950)%,NRE、PRE均高于我国植物叶片平均重吸收率,KRE高于全球植物平均重吸收率。叶片化学计量特征与PRE相关性较强,与NRE和KRE相关性稍差,狗尾草叶片养分重吸收率主要受衰老叶化学计量比的影响。研究区内狗尾草重吸收率较高,其化学计量比与养分重吸收存在一定相关性,说明狗尾草适应性极强,能够很好地适应贫瘠的环境,对海岸带防护林的养护有重要意义。
Abstract:We explored the adaptation of Setaria viridis (L.) Beauv. to harsh sandy coastal environments in the core area of the Yantai Coastal Shelter Forest Nature Reserve by measuring the carbon (C), nitrogen (N), phosphorus (P), and potassium (K) content in mature and senescent leaves, analyzing their ecological stoichiometric characteristics and nutrient resorption efficiencies, and studying the coupling relationship between the two. Results showed that leaf N∶P was greater than 16 and the growth and development of S. viridis was more restricted by P. Leaf nitrogen resorption efficiency (NRE), phosphorus resorption efficiency (PRE), and potassium resorption efficiency (KRE) were (58.038 ± 2.973)%, (62.167 ± 5.301)%, and (86.557 ± 0.950)%, respectively. NRE and PRE were higher than the average plant leaf resorption efficiency in China and KRE was higher than the global average plant resorption efficiency. The leaf stoichiometric characteristics were strongly correlated with PRE, but slightly less correlated with NRE and KRE. Nutrient resorption efficiency of the leaves was mainly affected by the stoichiometric ratio of the senescent leaves. The nutrient resorption efficiency of S. viridis was high in the study area, with a certain correlation between the stoichiometric ratio and nutrient resorption. These results indicate that S. viridis exhibits strong adaptability to the barren environment, which is of great significance to the conservation of coastal protection forests.
-
各种元素在植物体内是紧密联系、同步变化的。植物养分重吸收的过程不仅会引起某一种元素发生变化,还同时影响多种元素的变化[1],衰老叶片的养分重吸收与植物养分状况和生物地球化学循环密切相关。植物体内C、N、P、K元素含量及其生态化学计量特征在指示植物对环境变化的响应和适应性方面发挥着重要作用[2]。一般认为, C∶N∶P∶K的化学计量学具有内在的稳定性[3]。植物N∶P可以反映N或P是否限制植物生长[4,5],C∶N、C∶P则可以反映植物对所在环境的适应性[6]。生态化学计量特征从元素组成的角度探索 C、N、P、K等元素的生态过程和生物地球化学循环,是研究植物生长过程中养分限制情况以及植物与土壤之间元素循环的重要手段。
养分重吸收(NuR)是指衰老叶片将养分转移至成熟叶片中,使植物得以重复利用衰老叶片中残留营养元素的过程[7]。通过植物养分重吸收,叶片养分得以重复使用,而不是随着落叶而流失,促进植物生长、繁殖,提高植物竞争能力,特别是在营养贫乏的环境中,植物生长强烈依赖于衰老组织的养分重吸收[8]。植物养分重吸收率被广泛应用于评价植物对环境的适应性响应,营养限制对植物生长的影响以及生物多样性与初级生产力之间的相关性[9]。研究发现,养分重吸收具有地理格局,受温度、湿度、降水和土壤养分状况等非生物因素的影响[7]。其中,土壤养分状况与植物养分重吸收之间的关系一直饱受争议。一般认为,土壤养分状况与植物养分重吸收率呈负相关,植物在贫瘠环境下养分重吸收率较高[10]。但基于不同的研究可能会得出不同结论,因此土壤养分状况与植物养分重吸收率之间的关系还有待进一步研究。
狗尾草(Setaria viridis (L.) Beauv.)适应性极强,具有极强的耐贫瘠、耐干旱和耐盐碱等抗逆特性[11]。生长在烟台砂质海岸防护林下的狗尾草属于黑松(Pinus thunbergii Parl.)群落中草本层的优势种,不仅丰富了防护植被的生物多样性,而且在保持水土、降低土壤砂性、减缓土壤盐碱化进程等方面发挥重要作用[12]。目前,国内外关于植物重吸收的研究多集中于乔木和草地中的草本植物,关于森林生态系统中草本植物的研究相对较少,尤其是狗尾草分布面积较小,没有引起人们足够的重视,有关狗尾草养分重吸收的研究鲜有报道[13]。本文研究了狗尾草成熟叶和衰老叶的C、N、P、K化学计量比和N、P、K重吸收效率,分析狗尾草生长过程中的养分限制因子,研究了狗尾草养分重吸收与化学计量比之间的偶联关系,以探究狗尾草体内元素的变化规律和对恶劣环境的适应,为更好地利用植物特性促进烟台砂质海岸的恢复与建设提供理论参考。
1. 研究地区与研究方法
1.1 研究区概况
研究区位于山东省烟台市沿海防护林省级自然保护区核心区内(37°26′~37°28′ N,121°49′~121°55′ E),属暖温带东亚季风型大陆性气候,受季风和海洋影响显著[14],年均降水量为737.2 mm,年均气温11.5℃,年均相对湿度68%,年均日照时数2698.4 h,平均无霜期180 d[15]。
研究区地势低平,土壤为微酸性滨海砂土,有机质含量极低,土壤贫瘠(表1)。黑松防护林下主要草本植物为狗尾草、滨藜(Atriplex patens (Litv.) Iljin.)、美洲商陆(Phytolacca ameyicana L.)、砂引草(Messerschmidia sibirica L.)、狗牙根(Cynodon dactylon (L.) Pers.)、马齿苋(Portulaca oleracea L.)等[16]。
表 1 本研究区与其他地区土壤养分含量比较Table 1. Comparison of soil nutrient contents between this study area and other areas研究区域
Study area全氮TN /
g/kg全磷TP /
g/kg硝态氮NO3− -N /
mg/kg铵态氮NH4 + -N /
mg/kg速效磷AP /
mg/kg速效钾AK /
mg/kg有机碳SOC /
g/kg文献
Reference本研究区 0.263 0.060 2.740 4.831 15.516 9.220 8.006 本研究 科尔沁沙地灌丛内部 0.190 0.009 − − − − 3.740 [17] 科尔沁沙地灌丛外部 0.070 0.007 − − − − 2.620 [17] 中国北方温带草原 1.440 0.370 13.480 − 4.150 − 17.400 [18] 中国陆地土壤 24.560 1.800 − − − − 0.780 [19] 1.2 样品采集
2021年9月,以烟台沿海防护林省级自然保护区核心区段内的狗尾草为研究对象,在垂直于海岸线方向,以300 m为间隔平行设置3条样带,在各样带每隔200 m设置1个采样点,每条样带由沿海向内陆各设置7个样点,共计21个样点,分别对狗尾草成熟叶和衰老叶进行采样。
每个样点设置1个20 m × 20 m的样方,在其东北、西南对角线上各设置1个1 m × 1 m的草本样方。记录样方内全部狗尾草数量、高度、盖度、基径,选取10株长势良好的狗尾草为取样对象。选取颜色深绿、健康无病虫害、轮廓完整的叶片作为成熟叶片,选取颜色枯黄、健康无病虫害、自然衰老的叶片作为衰老叶片[20]。植物叶片样品经去离子水冲洗后,于105℃烘箱中杀青10 min,75℃烘干至恒重,冷却后采用多功能粉碎机磨碎后过0.25 mm筛,然后将样品编号并分别测定狗尾草成熟叶和衰老叶有机碳(Organic carbon, OC)、全氮(Total nitrogen, TN)、全磷(Total phosphorus, TP)、全钾(Total potassium, TK)的含量。
1.3 化学测量
植物样品经消煮后,采用重铬酸钾(K2Cr2O7)—氧化外加热法测定有机碳含量。经H2SO4-H2O2消解后,采用奈氏比色法测定全氮含量,钼锑抗比色法测定全磷含量,全钾含量则采用火焰光度法测定。
1.4 数据分析
叶片养分重吸收率(NuRE)采用Aerts[21]公式计算如下:
NuRE=(1−XSenescedXMature )×100% (1) 式中,NuRE为叶片N、P或K的重吸收效率,叶片N、P、K的养分重吸收率分别以NRE、PRE、KRE表示。XMature代表成熟叶片中N、P或K的含量(g/kg),XSenesced代表衰老叶片中N、P或K的含量(g/kg)。Van Heerwaarden等[22]和Luyssaert等[23]研究对比发现,基于叶面积进行的养分重吸收率的计算可能比基于叶质量进行的计算更准确一些,但在衰老过程中可能发生叶面积收缩,并最终导致重吸收效率的降低。狗尾草叶片在衰老期间显著收缩,因此,本研究仍使用基于质量的养分重吸收率,但没有考虑衰老过程中的叶片质量损失,可能会导致低估营养吸收。
运用Excel和SPSS19.0软件进行数据分析,利用ArcMap 10.2和Origin 2021软件绘图。狗尾草叶片生态化学计量比采用百分比表示,采用单因素ANOVA检验中的Duncan(D)法分析7个不同海岸距离下采样点叶片养分含量以及生态化学计量比之间的差异,并采用Pearson相关分析确定其叶片养分含量、化学计量比与叶片养分重吸收率之间的关系。P < 0.05表示显著相关,P < 0.01表示极显著相关。
2. 结果与分析
2.1 狗尾草叶片C、N、P、K含量变化特征
由表2可知,狗尾草成熟叶和衰老叶C、N、P、K含量存在差异。狗尾草成熟叶C含量在443.431~542.204 g/kg,N含量在17.107~20.224 g/kg,P含量在0.770~1.900 g/kg,K含量在14.204~21.822 g/kg;C、N、P、K平均含量分别为(516.236 ± 35.130)、(19.061 ± 1.216)、(1.218 ± 0.223)和(17.074 ± 2.734) g/kg。成熟叶C含量差异显著(P < 0.05),N、P、K含量无显著差异。
表 2 狗尾草叶片C、N、P、K元素含量Table 2. C, N, P, and K contents in Setaria viridis leaves海岸距离
Coastal
distance / m成熟叶 Mature leaves 衰老叶Senescent leaves C / g/kg N / g/kg P / g/kg K / g/kg C / g/kg N / g/kg P / g/kg K / g/kg 200 500.652 ± 18.164ab 17.107 ± 4.425a 1.900 ± 0.210a 16.771 ± 2.277a 468.712 ± 84.033a 5.287 ± 1.761c 1.445 ± 0.402a 2.064 ± 0.892a 400 542.204 ± 0.885a 18.706 ± 1.309a 1.146 ± 0.152a 16.895 ± 2.418a 522.466 ± 24.322a 7.359 ± 0.960bc 0.555 ± 0.100b 2.150 ± 0.464a 600 536.175 ± 12.554a 19.522 ± 1.047a 0.922 ± 0.315a 14.204 ± 1.246a 526.472 ± 52.701a 6.793 ± 1.143bc 0.253 ± 0.061b 2.062 ± 0.297a 800 530.873 ± 35.351a 17.826 ± 1.018a 0.812 ± 0.076a 15.210 ± 1.191a 521.339 ± 22.216a 8.136 ± 0.675ab 0.312 ± 0.136b 2.366 ± 0.177a 1000 539.835 ± 15.570a 20.224 ± 1.585a 0.770 ± 0.069a 15.031 ± 2.326a 532.086 ± 68.120a 7.286 ± 0.798bc 0.235 ± 0.058b 1.661 ± 0.436a 1200 520.482 ± 15.945a 20.003 ± 4.275a 1.438 ± 0.400a 19.586 ± 7.709a 527.750 ± 10.502a 9.616 ± 0.885a 0.644 ± 0.140b 2.585 ± 0.242a 1400 443.431 ± 78.507b 20.039 ± 4.671a 1.540 ± 0.386a 21.822 ± 7.187a 351.756 ± 8.377b 6.898 ± 0.951bc 0.934 ± 0.255ab 4.415 ± 0.449a 注:同列不同字母表示差异显著(P < 0.05)。下同。 Notes: Means that do not share a letter are significantly different (P < 0.05). Same below. 狗尾草衰老叶C含量在351.756~532.086 g/kg,N含量在5.287~9.616 g/kg,P含量在0.235~1.445 g/kg,K含量在1.661~4.415 g/kg,C、N、P、K平均含量分别为(492.940 ± 65.913)、(7.339 ± 1.325)、(0.626 ± 0.140)、(2.472 ± 0.903) g/kg。衰老叶C、N、P含量差异显著(P < 0.05),K含量无显著差异。
2.2 狗尾草成熟叶和衰老叶C、N、P、K生态化学计量特征
由表3可知,狗尾草成熟叶C∶N在23.667~31.052,C∶P在266.235~703.351,C∶K在22.296~37.888,N∶P在8.929~26.460,N∶K在0.957~1.380,K∶P在8.840~20.491。狗尾草衰老叶C∶N在55.179~81.934,C∶P在351.925~2355.906,C∶K在112.346~333.929,N∶P在5.007~32.762,N∶K在2.127~4.755,K∶P在2.193~8.309。成熟叶C∶N、C∶P、C∶K、N∶P、N∶K、K∶P平均值分别为27.445 ± 2.408、542.599 ± 57.955、31.436 ± 5.426、19.581 ± 5.647、1.154 ± 0.165、16.994 ± 4.104;衰老叶C∶N、C∶P、C∶K、N∶P、N∶K、K∶P平均值分别为66.437 ± 11.377、1368.538 ± 424.432、240.922 ± 68.672、20.784 ± 9.020、3.687 ± 1.091、5.827 ± 2.075。C∶N、C∶P、C∶K、N∶P、N∶K在衰老叶中比值较高,K∶P则相反,K∶P在成熟叶中比值较高。C∶N、C∶P、C∶K、N∶K、K∶P在成熟叶和衰老叶之间差异较大,N∶P在成熟叶和衰老叶之间差异较小。成熟叶C∶P、C∶K、N∶P、N∶K、K∶P差异显著,C∶N无显著差异。衰老叶C∶P、C∶K、N∶P、N∶K差异显著,C∶N、K∶P无显著差异。
表 3 狗尾草成熟叶和衰老叶C、N、P、K化学计量比Table 3. Stoichiometric ratios of C, N, P, and K in mature and senescent leaves of Setaria viridis叶类型
Type of leaf海岸距离
Coastal distance / mC∶N C∶P C∶K N∶P N∶K K∶P 成熟叶 200 31.052 ± 7.393a 266.235 ± 37.082b 30.357 ± 5.590ab 8.929 ± 1.649b 1.009 ± 0.149ab 8.840 ± 0.962b 400 26.529 ± 6.326a 613.489 ± 174.042ab 29.083 ± 4.102ab 21.477 ± 6.405a 1.125 ± 0.205ab 20.491 ± 4.816a 600 27.521 ± 1.675a 632.747 ± 131.660ab 37.888 ± 2.418a 22.835 ± 7.343a 1.380 ± 0.121a 16.531 ± 5.214ab 800 29.832 ± 2.340a 655.602 ± 40.326ab 35.119 ± 4.450a 22.020 ± 1.266a 1.175 ± 0.074ab 18.842 ± 2.278ab 1000 26.823 ± 2.543a 703.351 ± 42.353a 36.410 ± 4.940a 26.460 ± 3.960a 1.364 ± 0.200a 19.487 ± 2.038ab 1200 26.691 ± 4.804a 449.660 ± 74.570ab 28.898 ± 8.884ab 16.424 ± 6.709ab 1.071 ± 0.222ab 14.916 ± 3.200ab 1400 23.667 ± 5.194a 477.108 ± 113.887ab 22.296 ± 7.187b 18.921 ± 5.130ab 0.957 ± 0.291b 19.850 ± 4.496ab 衰老叶 200 81.934 ± 41.860a 351.925 ± 71.100b 209.155 ± 78.842ab 5.007 ± 1.298b 2.430 ± 0.034bc 2.193 ± 0.125a 400 59.987 ± 22.376a 1447.328 ± 325.356ab 232.800 ± 76.836ab 22.152 ± 5.590ab 3.954 ± 0.653abc 5.913 ± 1.273a 600 79.024 ± 13.599a 2159.777 ± 481.354a 259.182 ± 39.733ab 27.194 ± 2.466a 3.290 ± 0.182abc 8.309 ± 1.188a 800 59.313 ± 6.926a 1451.048 ± 184.236ab 275.393 ± 85.796ab 24.341 ± 4.735ab 4.625 ± 1.154ab 5.041 ± 1.385a 1000 3.580 ± 7.552a 2355.906 ± 568.452a 333.930 ± 79.235a 32.762 ± 7.167a 4.628 ± 1.532ab 7.274 ± 2.223a 1200 55.179 ± 4.911a 1100.669 ± 260.227ab 263.649 ± 99.345ab 19.444 ± 4.797ab 4.755 ± 1.580a 4.674 ± 0.842a 1400 56.046 ± 10.468a 713.111 ± 75.989b 112.346 ± 35.597b 14.587 ± 2.333ab 2.127 ± 0.298c 7.383 ± 2.869a 2.3 狗尾草叶片N、P、K养分重吸收特征
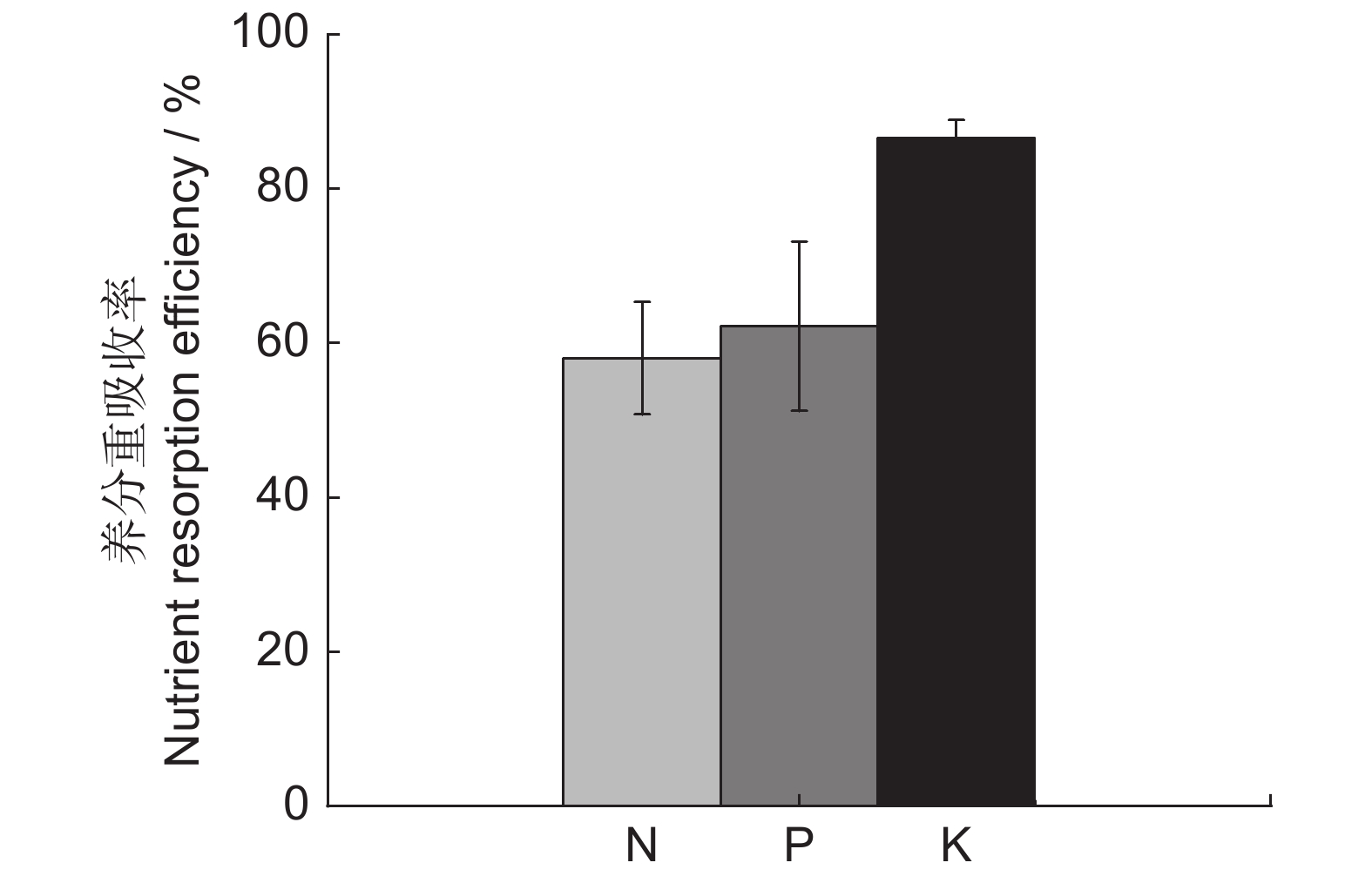
狗尾草叶片N、P、K养分重吸收率差异较大。NRE在46.700%~69.687%,PRE在34.003%~77.769%,KRE在81.545%~88.651%;NRE、PRE、KRE变异系数分别为5.123%、8.527%、1.098%,KRE变化幅度很小,比较稳定。狗尾草叶片的平均NRE、PRE、KRE分别为(58.038 ± 2.973)%、(62.167 ± 5.301)%、(86.557 ± 0.950)%,KRE > PRE > NRE,N、P、K养分重吸收效率均较高(图1)。
![]() 图 1 狗尾草叶片N、P、K养分重吸收率Figure 1. Nutrient resorption efficiency of N, P, and K in Setaria viridis
图 1 狗尾草叶片N、P、K养分重吸收率Figure 1. Nutrient resorption efficiency of N, P, and K in Setaria viridis2.4 狗尾草叶片C、N、P、K元素含量及化学计量比与重吸收率的相关性分析
本研究发现(表4、表5),狗尾草叶片C、N、P、K元素含量及化学计量比与重吸收率具有一定的相关性。狗尾草叶片NRE与成熟叶N含量呈极显著正相关(P < 0.01),与衰老叶N含量呈极显著负相关。PRE与成熟叶P含量相关性不显著,与衰老叶P含量呈极显著负相关。KRE与成熟叶N含量呈显著正相关,与成熟叶K含量相关性不显著,与衰老叶C含量呈显著正相关,与衰老叶K含量呈极显著负相关。
表 4 NRE、PRE、KRE与叶片C、N、P、K含量的相关性Table 4. Correlations among NRE, PRE, KRE, and C, N, P, and K contents in leaves指标
Index成熟叶 Mature leaves 衰老叶 Senescent leaves C N P K C N P K NRE −0.183 0.561** 0.019 0.291 −0.212 −0.762** 0.163 −0.023 PRE 0.155 0.198 −0.354 −0.273 0.209 0.162 −0.776** −0.362 KRE −0.049 0.513* 0.123 0.205 0.467* 0.168 −0.259 −0.961** Notes: *, P < 0.05; **, P < 0.01. Same below. 表 5 NRE、PRE、KRE与叶片C、N、P、K化学计量比的相关性Table 5. Correlations among NRE, PRE, KRE, and stoichiometric ratios of C, N, P, and K in leaves指标
Index成熟叶 Mature leaves 衰老叶 Senescent leaves C∶N C∶P C∶K N∶P N∶K K∶P C∶N C∶P C∶K N∶P N∶K K∶P NRE −0.571** 0.042 −0.291 0.217 0.145 0.175 0.580** 0.140 0.160 −0.009 −0.247 0.125 PRE −0.090 0.294 0.262 0.403 0.484* 0.220 −0.075 0.742** 0.213 0.808** 0.282 0.740** KRE −0.425 −0.107 −0.113 0.083 0.206 −0.096 0.169 0.319 0.738** 0.317 0.707** −0.208 狗尾草叶片NRE与成熟叶C∶N呈极显著负相关,与衰老叶C∶N呈极显著正相关。叶片PRE与成熟叶N∶K呈显著正相关 (P < 0.05),与衰老叶C∶P、N∶P、K∶P呈极显著正相关。叶片KRE与衰老叶C∶K、N∶K呈极显著正相关。成熟叶C∶P、C∶K、N∶P、K∶P与叶片 NRE、PRE、KRE的相关性均不显著。
3. 讨论
3.1 狗尾草叶片养分化学计量特征
植物养分浓度和化学计量比已被广泛用于表明植物吸收N、P、K元素时同化C的能力,即养分利用效率,并揭示哪些因素限制了植物生长和陆地净初级生产力[2]。本研究中狗尾草成熟叶C含量平均值为(516.236 ± 35.130) g/kg,高于Elser等[24]观测的全球植物平均C含量(464.000 g/kg),说明研究区狗尾草有机碳含量较高。成熟叶N、P含量平均值分别为(19.061 ± 1.216)和(1.218 ± 0.223) g/kg,均低于全球植物叶片平均N、P含量(分别为20.600、1.490 g/kg),也低于Han等[25]研究得出的中国陆地植物叶片平均N、P含量(分别为20.200、1.450 g/kg),但高于内蒙古草地植物平均N、P含量(分别为17.150、1.100 g/kg)[26],而与鄱阳湖湿地的苔草(Carex tristachya L.)和南荻(Triarrhena lutarioriparia (L.) Liu)[27]以及互花米草(Spartina alterniflora Lois.)[28]相比,C、N含量明显偏高,P含量略低,而与黄土丘陵区草本植物成熟叶(平均C、N、P、K含量分别为496.200、25.360、2.180、15.400 g/kg)[4]相比,C、K含量偏高,N、P含量偏低,这可能是因为不同植物对养分的利用不同。与科尔沁沙地灌丛内部、外部狗尾草N、P含量(灌丛内、外部狗尾草N、P含量分别为28.190、0.150 g/kg和17.260、0.230 g/kg)相比[17],除灌丛内部狗尾草N含量高于研究区狗尾草外,其余均低于研究区狗尾草成熟叶N、P含量。这可能是因为研究区土壤全氮、全磷含量均高于科尔沁沙地灌丛内、外部土壤含量,草本植物所需的营养元素主要来源于土壤[29],而不同地区土壤养分状况、土壤元素可利用率以及植物生长环境不同。
狗尾草衰老叶C、N、P、K元素含量均低于成熟叶,这是狗尾草叶片对C、N、P、K养分重吸收的结果,说明研究区狗尾草可以通过降低衰老叶中的养分含量,保存养分,促进养分回流,更好地适应营养贫乏的环境。衰老叶平均N、P含量分别为(7.339 ± 1.325)、(0.626 ± 0.140) g/kg,均低于全球植物衰老叶平均水平(10.000 、0.700 g/kg)[30],与成熟叶N、P含量相同,与黄土丘陵区草本植物衰老叶(平均C、N、P、K含量分别为409.220、11.410、1.690、3.230 g/kg)相比,C含量偏高,N、P、K含量偏低,也与成熟叶相似[4]。可能是由于衰老叶养分含量随成熟叶养分含量变动引起,衰老叶元素含量的一个重要决定因素是成熟叶元素含量。成熟叶C、N、P、K含量较低时,衰老叶的养分含量也相应较低。
叶片的C∶N、C∶P、C∶K可以表示植物同化碳元素的能力,也反映植物的固碳能力[31]。狗尾草衰老叶元素化学计量比较成熟叶高,体现了养分重吸收的结果。受叶片N 含量较低的影响,狗尾草叶片C∶N均高于全球植物平均C∶N(23.400)[32],也明显高于科尔沁沙地主要植物C∶N、C∶P(分别为17.800、228.600)[33],较高的 C∶N、C∶P反映了狗尾草N、P元素利用效率较高,表明狗尾草在养分贫瘠的砂质土壤中具有养分利用和固碳的优势[34]。N∶P的变化提供了对各种空间尺度上养分循环和限制的洞察,补充了植物养分生物地球化学的其他指标[35]。狗尾草成熟叶和衰老叶N∶P分别为19.581 ± 5.647和20.784 ± 9.020,均高于中国陆地植物叶片平均N∶P(16.300)[25],也高于内蒙古草地植物平均N∶P(17.900)[26],但低于鄱阳湖湿地优势植物N∶P(21.240)[27],可能是因为研究区位于海陆交接的滨海砂地,与陆地、草地、湿地相比生态系统具有独特性[36]。
植物叶片的N∶P表示N或P是否限制植物生长[37],Koerselman和Meulema[38]认为,N∶P > 16时,表示植物生长主要受到P元素限制,而N∶P < 14时,表示受到N元素限制。当N∶P在14 ~ 16时,N或P可以限制或植物生长受N和P共同限制。本研究中狗尾草N∶P > 16,表明该区域的狗尾草生长主要受到了P元素的限制,这与Han等[25]研究得出的的中国大多数土壤中植物可能受P限制一致,也与芦苇(Phragmites australis (Cav.) Trin. ex Steud.)[20]限制元素相同,但异于鄱阳湖湿地优势植物[27]。由Yuan等[39]的研究可知,温度和降水对生物地球化学的影响可能会导致温带土壤中N含量充足但P含量有限,且研究区域土壤为砂土,土壤贫瘠,总体肥力不足,可以通过施加有机肥提高土壤肥力。
3.2 狗尾草叶片养分重吸收特点
植物重吸收被认为是受自然选择影响的营养保护性状,有助于满足组织再生、化学成分平衡和生物量生产的需求[40]。狗尾草叶片NRE、PRE、KRE平均值分别为(58.038 ± 2.973)%、(62.167 ± 5.301)%和(86.557 ± 0.950)%,与Vergutz等[41]研究得出的全球植物NRE、PRE、KRE(分别为62.100%、64.900%、70.100%)相比,NRE、PRE均低于全球植物平均水平,KRE高于全球植物平均水平,但NRE、PRE均高于我国植物平均重吸收率(NRE、PRE分别为47.980%和54.380%)[42]。研究区土壤贫瘠,根据全国第2次土壤普查养分分级标准,研究区土壤有效氮含量属于6级(极缺)、速效磷含量属于3级(中)、速效钾含量属于6级(极缺),但KRE > PRE > NRE,PRE、KRE对土壤养分含量的响应较明显,NRE对土壤养分含量的响应不明显。狗尾草叶片NRE、PRE低于敦煌阳关湿地芦苇叶片(N、P重吸收效率分别为62.500%~76.100%、80.100%~84.500%)[20],但高于古尔班通古特沙漠中4种荒漠草本植物叶片平均N、P 重吸收效率(42.700%和54.700%)[43],也高于厚荚相思(Acacia crassicarpa A. Cunn Ex Benth.)(NRE、PRE分别为27.160%、58. 080%)[44]和5龄、11龄枯黄期青海扁茎早熟禾(Poa pratensis var. anceps Gaud.)(5龄青海扁茎早熟禾NRE、PRE分别为30.180%、29.420%,11龄青海扁茎早熟禾NRE、PRE为28.380%、28.770%)[45]。这可能与不同研究区土壤养分含量以及测量方法不同有关。Vergutz等[41]研究发现当使用基于叶质量的浓度时,叶片质量损失可能导致平均重吸收率低估10%,而使用基于面积的浓度时,叶片收缩可能导致平均低估6%。
狗尾草叶片KRE均高于NRE、PRE,这说明狗尾草对环境中的K元素依赖性较弱,而对N、P元素依赖性较强,这与王振南等[1]对苜蓿(Medicago sativa L.)叶片的研究结果一致,也与Vergutz等[41]研究得出的禾本科植物具有较高的KRE相吻合。狗尾草叶片PRE高于NRE,这可能是由于植物从土壤中获取P元素比从土壤中获取N元素少,或者从衰老叶片中吸收P元素比从土壤中吸收P元素更容易,狗尾草需要提高PRE来降低对土壤中磷元素的需求[46]。此外,根据相对再吸收假说[47],PRE-NRE > 0即PRE较高时,表明狗尾草的生长受到P元素的限制,对应于狗尾草叶片N∶P > 16,进一步证明了研究区内狗尾草生长主要受到了P元素的限制。
狗尾草叶片NRE、PRE、KRE分别与衰老叶N、P、K元素呈极显著负相关,适应营养贫乏环境的植物具有较高的重吸收效率和较低的衰老叶片养分含量,说明叶片养分重吸收促进了衰老叶养分的重新分配利用。狗尾草叶片NRE与成熟叶N元素呈极显著正相关,KRE与成熟叶K元素呈显著正相关,说明成熟叶中较高的N、K含量可以促进养分重吸收。狗尾草叶片PRE与成熟叶P元素呈负相关,这反映了植物在从土壤中吸收养分和从衰老叶中吸收养分之间的相对成本权衡[48]。Liu 等[48]研究发现,如果植物从养分贫瘠的土壤中吸收养分比从衰老叶中吸收养分成本更高,那么重吸收率就会随着成熟叶养分含量的降低而增加。研究区土壤磷含量缺乏,狗尾草生长主要受P元素限制,所以可能从衰老叶中吸收P元素更容易一些。NRE和PRE的相反模式可以反映它们不同的生化作用,可能是狗尾草为了平衡体内元素含量,触发P限制下的特殊适应机制[49]。狗尾草叶片养分重吸收效率主要受衰老叶养分含量的影响,且为负向影响,其次受成熟叶养分含量的正向影响,这与刘冬等[20]和 Kobe等[50]的研究结果相反,但与王振南等[1]和Wang等[18]的研究一致,植物种类、生理特性、气候土壤条件以及植物整体营养水平不同等都是造成差别的原因。
3.3 狗尾草叶片养分重吸收与化学计量比的偶联关系
植物生长不仅依赖于养分的有效性,还依赖于多种养分之间的平衡[51]。狗尾草叶片化学计量比和重吸收效率是叶片C、N、P、K元素综合作用的产物,所以化学计量比和重吸收效率之间必然存在一定的偶联关系[1]。狗尾草叶片养分重吸收率与成熟叶、衰老叶化学计量比均主要呈正相关,说明化学计量比所指示的相对养分限制可以很好地预测狗尾草叶片N、P、K 的重吸收效率[48, 51]。狗尾草叶片重吸收率与成熟叶化学计量比相关性稍弱,与衰老叶化学计量比相关性显著,这表明衰老叶化学计量比对狗尾草叶片养分重吸收效率的影响更大,与养分重吸收率主要受衰老叶养分含量影响相对应。
狗尾草成熟叶、衰老叶化学计量特征与PRE相关性较强,与NRE和KRE相关性稍差。这可能是因为PRE与NRE的主要控制因素不同。Tang等[52]研究发现,NRE与生物因素的相关性高于非生物因素,而PRE与非生物因素的相关性高于生物因素。N是一种更来源于生物的营养物质,而P是一种更来源于岩石的营养物质。狗尾草在土壤磷含量缺乏的情况下,主要依靠养分重吸收从衰老叶中吸收P元素来满足自身发展的需要,所以PRE与化学计量特征相关性较强。目前,关于植物养分重吸收率与化学计量比之间偶联关系的研究还比较少,相关研究多集中在分别研究植物养分重吸收率、化学计量比的特征,因此,要深入了解二者之间的偶联关系还需要进一步开展定量研究。
-
![]()
图 1 狗尾草叶片N、P、K养分重吸收率
Figure 1. Nutrient resorption efficiency of N, P, and K in Setaria viridis
表 1 本研究区与其他地区土壤养分含量比较
Table 1 Comparison of soil nutrient contents between this study area and other areas
研究区域
Study area全氮TN /
g/kg全磷TP /
g/kg硝态氮NO3− -N /
mg/kg铵态氮NH4 + -N /
mg/kg速效磷AP /
mg/kg速效钾AK /
mg/kg有机碳SOC /
g/kg文献
Reference本研究区 0.263 0.060 2.740 4.831 15.516 9.220 8.006 本研究 科尔沁沙地灌丛内部 0.190 0.009 − − − − 3.740 [17] 科尔沁沙地灌丛外部 0.070 0.007 − − − − 2.620 [17] 中国北方温带草原 1.440 0.370 13.480 − 4.150 − 17.400 [18] 中国陆地土壤 24.560 1.800 − − − − 0.780 [19]  下载: 导出CSV
下载: 导出CSV
表 2 狗尾草叶片C、N、P、K元素含量
Table 2 C, N, P, and K contents in Setaria viridis leaves
海岸距离
Coastal
distance / m成熟叶 Mature leaves 衰老叶Senescent leaves C / g/kg N / g/kg P / g/kg K / g/kg C / g/kg N / g/kg P / g/kg K / g/kg 200 500.652 ± 18.164ab 17.107 ± 4.425a 1.900 ± 0.210a 16.771 ± 2.277a 468.712 ± 84.033a 5.287 ± 1.761c 1.445 ± 0.402a 2.064 ± 0.892a 400 542.204 ± 0.885a 18.706 ± 1.309a 1.146 ± 0.152a 16.895 ± 2.418a 522.466 ± 24.322a 7.359 ± 0.960bc 0.555 ± 0.100b 2.150 ± 0.464a 600 536.175 ± 12.554a 19.522 ± 1.047a 0.922 ± 0.315a 14.204 ± 1.246a 526.472 ± 52.701a 6.793 ± 1.143bc 0.253 ± 0.061b 2.062 ± 0.297a 800 530.873 ± 35.351a 17.826 ± 1.018a 0.812 ± 0.076a 15.210 ± 1.191a 521.339 ± 22.216a 8.136 ± 0.675ab 0.312 ± 0.136b 2.366 ± 0.177a 1000 539.835 ± 15.570a 20.224 ± 1.585a 0.770 ± 0.069a 15.031 ± 2.326a 532.086 ± 68.120a 7.286 ± 0.798bc 0.235 ± 0.058b 1.661 ± 0.436a 1200 520.482 ± 15.945a 20.003 ± 4.275a 1.438 ± 0.400a 19.586 ± 7.709a 527.750 ± 10.502a 9.616 ± 0.885a 0.644 ± 0.140b 2.585 ± 0.242a 1400 443.431 ± 78.507b 20.039 ± 4.671a 1.540 ± 0.386a 21.822 ± 7.187a 351.756 ± 8.377b 6.898 ± 0.951bc 0.934 ± 0.255ab 4.415 ± 0.449a 注:同列不同字母表示差异显著(P < 0.05)。下同。 Notes: Means that do not share a letter are significantly different (P < 0.05). Same below.
下载: 导出CSV
表 3 狗尾草成熟叶和衰老叶C、N、P、K化学计量比
Table 3 Stoichiometric ratios of C, N, P, and K in mature and senescent leaves of Setaria viridis
叶类型
Type of leaf海岸距离
Coastal distance / mC∶N C∶P C∶K N∶P N∶K K∶P 成熟叶 200 31.052 ± 7.393a 266.235 ± 37.082b 30.357 ± 5.590ab 8.929 ± 1.649b 1.009 ± 0.149ab 8.840 ± 0.962b 400 26.529 ± 6.326a 613.489 ± 174.042ab 29.083 ± 4.102ab 21.477 ± 6.405a 1.125 ± 0.205ab 20.491 ± 4.816a 600 27.521 ± 1.675a 632.747 ± 131.660ab 37.888 ± 2.418a 22.835 ± 7.343a 1.380 ± 0.121a 16.531 ± 5.214ab 800 29.832 ± 2.340a 655.602 ± 40.326ab 35.119 ± 4.450a 22.020 ± 1.266a 1.175 ± 0.074ab 18.842 ± 2.278ab 1000 26.823 ± 2.543a 703.351 ± 42.353a 36.410 ± 4.940a 26.460 ± 3.960a 1.364 ± 0.200a 19.487 ± 2.038ab 1200 26.691 ± 4.804a 449.660 ± 74.570ab 28.898 ± 8.884ab 16.424 ± 6.709ab 1.071 ± 0.222ab 14.916 ± 3.200ab 1400 23.667 ± 5.194a 477.108 ± 113.887ab 22.296 ± 7.187b 18.921 ± 5.130ab 0.957 ± 0.291b 19.850 ± 4.496ab 衰老叶 200 81.934 ± 41.860a 351.925 ± 71.100b 209.155 ± 78.842ab 5.007 ± 1.298b 2.430 ± 0.034bc 2.193 ± 0.125a 400 59.987 ± 22.376a 1447.328 ± 325.356ab 232.800 ± 76.836ab 22.152 ± 5.590ab 3.954 ± 0.653abc 5.913 ± 1.273a 600 79.024 ± 13.599a 2159.777 ± 481.354a 259.182 ± 39.733ab 27.194 ± 2.466a 3.290 ± 0.182abc 8.309 ± 1.188a 800 59.313 ± 6.926a 1451.048 ± 184.236ab 275.393 ± 85.796ab 24.341 ± 4.735ab 4.625 ± 1.154ab 5.041 ± 1.385a 1000 3.580 ± 7.552a 2355.906 ± 568.452a 333.930 ± 79.235a 32.762 ± 7.167a 4.628 ± 1.532ab 7.274 ± 2.223a 1200 55.179 ± 4.911a 1100.669 ± 260.227ab 263.649 ± 99.345ab 19.444 ± 4.797ab 4.755 ± 1.580a 4.674 ± 0.842a 1400 56.046 ± 10.468a 713.111 ± 75.989b 112.346 ± 35.597b 14.587 ± 2.333ab 2.127 ± 0.298c 7.383 ± 2.869a
下载: 导出CSV
表 4 NRE、PRE、KRE与叶片C、N、P、K含量的相关性
Table 4 Correlations among NRE, PRE, KRE, and C, N, P, and K contents in leaves
指标
Index成熟叶 Mature leaves 衰老叶 Senescent leaves C N P K C N P K NRE −0.183 0.561** 0.019 0.291 −0.212 −0.762** 0.163 −0.023 PRE 0.155 0.198 −0.354 −0.273 0.209 0.162 −0.776** −0.362 KRE −0.049 0.513* 0.123 0.205 0.467* 0.168 −0.259 −0.961** Notes: *, P < 0.05; **, P < 0.01. Same below.
下载: 导出CSV
表 5 NRE、PRE、KRE与叶片C、N、P、K化学计量比的相关性
Table 5 Correlations among NRE, PRE, KRE, and stoichiometric ratios of C, N, P, and K in leaves
指标
Index成熟叶 Mature leaves 衰老叶 Senescent leaves C∶N C∶P C∶K N∶P N∶K K∶P C∶N C∶P C∶K N∶P N∶K K∶P NRE −0.571** 0.042 −0.291 0.217 0.145 0.175 0.580** 0.140 0.160 −0.009 −0.247 0.125 PRE −0.090 0.294 0.262 0.403 0.484* 0.220 −0.075 0.742** 0.213 0.808** 0.282 0.740** KRE −0.425 −0.107 −0.113 0.083 0.206 −0.096 0.169 0.319 0.738** 0.317 0.707** −0.208
下载: 导出CSV
-
[1] 王振南,赵梅,杨燕,李富宽,王慧,吕慎金. 苜蓿叶片氮、磷和钾养分重吸收与化学计量比的偶联关系[J]. 草业学报,2019,28(11):177−183. Wang ZN,Zhao M,Yang Y,Li FK,Wang H,Lü SJ. Relationships between alfalfa leaf nutrient resorption and stoichiometric ratios of nitrogen,phosphorus,and potassium[J]. Acta Prataculturae Sinica,2019,28 (11):177−183.
[2] Tong R,Zhou BZ,Jiang LN,Ge XG,Cao YH. Spatial patterns of leaf carbon,nitrogen,and phosphorus stoichiometry and nutrient resorption in Chinese fir across subtropical China[J]. Catena,2021,201:105221. doi: 10.1016/j.catena.2021.105221
[3] 阿里木·买买提,李翔,卡哈尔曼·恰依扎旦,常顺利. 天山云杉林下优势草本植物化学计量内稳性特征[J]. 西北林学院学报,2022,37(2):68−74. Alim·Mamat,Li X,Kahlman·Chayizadan,Chang SL. Nutrient and stoichiometric characteristics of dominant herbaceous in spruce forests on the northern slope of Tianshan mountains[J]. Journal of Northwest Forestry University,2022,37 (2):68−74.
[4] 刘洋,曾全超,安韶山,李鑫,黄懿梅. 黄土丘陵区草本植物叶片与枯落物生态化学计量学特征[J]. 应用生态学报,2017,28(6):1793−1800. Liu Y,Zeng QC,An SS,Li X,Huang YM. Ecological stoichiometry of leaf and litter of herbaceous plants in loess hilly and gully regions,China[J]. Chinese Journal of Applied Ecology,2017,28 (6):1793−1800.
[5] 王凯,沈潮,孙冰,王潇楠,魏东,吕林有. 干旱胁迫对科尔沁沙地榆树幼苗C、N、P化学计量特征的影响[J]. 应用生态学报,2018,29(7):2286−2294. Wang K,Shen C,Sun B,Wang XN,Wei D,Lü LY. Effects of drought stress on C,N and P stoichiometry of Ulmus pumila seedlings in Horqinsandy land,China[J]. Chinese Journal of Applied Ecology,2018,29 (7):2286−2294.
[6] Yuan ZY,Chen HYH,Reich PB. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus[J]. Nat Commun,2011,2 (1):344. doi: 10.1038/ncomms1346
[7] 周丽丽,钱瑞玲,李树斌,董博微,陈宝英,潘辉. 滨海沙地主要造林树种叶片功能性状及养分重吸收特征[J]. 应用生态学报,2019,30(7):2320−2328. Zhou LL,Qian RL,Li SB,Dong BW,Chen BY,Pan H. Leaf functional traits and nutrient resorption among major silviculture tree species in coastal sandy site[J]. Chinese Journal of Applied Ecology,2019,30 (7):2320−2328.
[8] Li YL,Jing C,Mao W,Cui D,Wang XY,Zhao XY. N and P resorption in a pioneer shrub (Artemisia halodendron) inhabiting severely desertified lands of Northern China[J]. J Arid Land,2014,6 (2):174−185. doi: 10.1007/s40333-013-0222-7
[9] Su Y,Ma XF,Gong YM,Li KH,Han WX,Liu XJ. Responses and drivers of leaf nutrients and resorption to nitrogen enrichment across northern China’s grasslands:a meta-analysis[J]. Catena,2021,199:105110. doi: 10.1016/j.catena.2020.105110
[10] 张效境,梁潇洒,马望,王正文. 呼伦贝尔草地植物茎秆和叶片中养分的时间动态与回收[J]. 植物生态学报,2021,45(7):738−748. doi: 10.17521/cjpe.2021.0125 Zhang XJ,Liang XS,Ma W,Wang ZW. Temporal variation and resorption of nutrients in plant culms and leaves in Hulun Buir grassland[J]. Chinese Journal of Plant Ecology,2021,45 (7):738−748. doi: 10.17521/cjpe.2021.0125
[11] 吴子龙,王磊,张浩,王晓改,李丹,赵昕. 狗尾草应用基础的研究进展[J]. 北方园艺,2021(5):130−135. Wu ZL,Wang L,Zhang H,Wang XG,Li D,Zhao X. Research progress on application basis of Setaria viridis[J]. Northern Horticulture,2021 (5):130−135.
[12] 骆娟,耿静,王宏信. 5种滨海沙生植物氮磷钾化学计量特征分析[J]. 现代农业科技,2020(10):144−145. Luo J,Geng J,Wang HX. Analysis on stoichiometric characteristics of N,P,K of five coastal sandy plants[J]. Modern Agricultural Science and Technology,2020 (10):144−145.
[13] 张崇邦,李钧敏. 退化羊草草原金狗尾草的抗盐碱生理及其生长生态特性的研究[J]. 哈尔滨师范大学自然科学学报,2001,17(2):93−97. Zhang CB,Li JM. The study on anti-salty and alkali physiology and growth ecological characteristics of Setaria lutescens on degenerative Leymus chinensis grassland[J]. Natural Science Journal of Harbin Normal University,2001,17 (2):93−97.
[14] 王琼. 牟平海岸带土壤理化性质分析[J]. 安徽农业科学,2015,43(28):95−97. Wang Q. Analysis of soil physical and chemical properties in coastal zone in Muping[J]. Journal of Anhui Agricultural Sciences,2015,43 (28):95−97.
[15] 韩广轩,王光美,张志东,李秋艳,薛钦昭. 烟台海岸黑松防护林种群结构及其随离岸距离的变化[J]. 林业科学,2008,44(10):8−13. doi: 10.3321/j.issn:1001-7488.2008.10.002 Han GX,Wang GM,Zhang ZD,Li QY,Xue QZ. Population structure of the Pinus thunbergii coastal protection forest and its spatial variation at different distances to coastline in Yantai[J]. Scientia Silvae Sinicae,2008,44 (10):8−13. doi: 10.3321/j.issn:1001-7488.2008.10.002
[16] 刘艳莉,陈鹏东,侯玉平,卜庆梅,柏新富. 烟台沙质海岸前沿4种草本植物热值与建成成本分析[J]. 生态环境学报,2018,27(7):1211−1217. Liu YL,Chen PD,Hou YP,Bu QM,Bai XF. Calorific value and construction cost of 4 herbaceous species in the coastal frontier of Yantai[J]. Ecology and Environmental Sciences,2018,27 (7):1211−1217.
[17] 毛伟,李玉霖,赵学勇,黄迎新,王少昆. 科尔沁沙地灌丛内外草本植物狗尾草叶性状的比较研究[J]. 草业学报,2009,18(6):144−150. doi: 10.11686/cyxb20090619 Mao W,Li YL,Zhao XY,Huang YX,Wang SK. Effect of Caragana microphylla on leaf traits of Setarria viridis in Horqin sandy land[J]. Acta Prataculturae Sinica,2009,18 (6):144−150. doi: 10.11686/cyxb20090619
[18] Wang Z,Jimoh SO,Li XL,Ji BM,Struik PC,et al. Different responses of plant N and P resorption to overgrazing in three dominant species in a typical steppe of Inner Mongolia,China[J]. PeerJ,2020,8:e9915. doi: 10.7717/peerj.9915
[19] Tian HQ,Chen GS,Zhang C,Melillo JM,Hall CAS. Pattern and variation of C∶N∶P ratios in China’s soils:a synthesis of observational data[J]. Biogeochemistry,2010,98 (1-3):139−151. doi: 10.1007/s10533-009-9382-0
[20] 刘冬,张剑,包雅兰,赵海燕,齐璇璇,等. 敦煌阳关湿地芦苇叶片养分重吸收模式及其对土壤水分的响应[J]. 应用生态学报,2020,31(3):807−813. Liu D,Zhang J,Bao YL,Zhao HY,Qi XX,et al. Nutrient resorption patterns of Phragmites australis leaves and its response to soil moisture in Yangguan wetland,Dunhuang,Northwest China[J]. Chinese Journal of Applied Ecology,2020,31 (3):807−813.
[21] Aerts R. Nutrient resorption from senescing leaves of perennials:are there general patterns?[J]. J Ecol,1996,84 (4):597−608. doi: 10.2307/2261481
[22] Van Heerwaarden LM,Toet S,Aerts R. Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization[J]. J Ecol,2003,91 (6):1060−1070. doi: 10.1046/j.1365-2745.2003.00828.x
[23] Luyssaert S,Staelens J,de Schrijver A. Does the commonly used estimator of nutrient resorption in tree foliage actually measure what it claims to?[J]. Oecologia,2005,144 (2):177−186. doi: 10.1007/s00442-005-0085-5
[24] Elser JJ,Fagan WF,Denno RF,Dobberfuhl DR,Folarin A,et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature,2000,408 (6812):578−580. doi: 10.1038/35046058
[25] Han WX,Fang JY,Guo DL,Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytol,2005,168 (2):377−385. doi: 10.1111/j.1469-8137.2005.01530.x
[26] 张良侠,樊江文,张文彦,钟华平. 内蒙古草地植物叶片氮、磷元素化学计量学特征分析[J]. 中国草地学报,2014,36(2):43−48. Zhang LX,Fan JW,Zhang WY,Zhong HP. Stoichiometry of leaf nitrogen and phosphorus in plants in grasslands in inner Mongolia[J]. Chinese Journal of Grassland,2014,36 (2):43−48.
[27] 郑艳明,尧波,吴琴,胡斌华,胡启武. 鄱阳湖湿地两种优势植物叶片C、N、P动态特征[J]. 生态学报,2013,33(20):6488−6496. doi: 10.5846/stxb201212041740 Zheng YM,Yao B,Wu Q,Hu BH,Hu QW. Dynamics of leaf carbon,nitrogen and phosphorus of two dominant species in a Poyang Lake wetland[J]. Acta Ecologica Sinica,2013,33 (20):6488−6496. doi: 10.5846/stxb201212041740
[28] 范全城,谢文霞,王志强,李萍. 互花米草碳、氮、磷化学计量特征及季节变化[J]. 环境科学与技术,2019,42(4):12−19. Fan QC,Xie WX,Wang ZQ,Li P. Seasonal variations in C,N and P stoichiometry of Spartina alterniflora[J]. Environmental Science & Technology,2019,42 (4):12−19.
[29] 刘艳艳. 武夷山自然保护区不同海拔甜槠林细根结构与养分的关系研究[D]. 福州: 福建农林大学, 2014. [30] Yuan ZY,Chen HYH. Global trends in senesced-leaf nitrogen and phosphorus[J]. Global Ecol Biogeogr,2009,18 (5):532−542. doi: 10.1111/j.1466-8238.2009.00474.x
[31] 周佳佳,罗有发,马李蓉,邢容容,吴永贵. 黔西北土法炼锌废渣堆场修复植物叶片和废渣的生态化学计量特征[J]. 地球与环境,2019,47(4):419−428. Zhou JJ,Luo YF,Ma LR,Xing RR,Wu YG. Ecological stoichiometric characteristics of plant leaves and slag at a restoration site of indigenous zinc smelting slag in northwestern Guizhou[J]. Earth and Environment,2019,47 (4):419−428.
[32] Ren CJ,Zhao FZ,Kang D,Yang GH,Han XH,et al. Linkages of C∶N∶P stoichiometry and bacterial community in soil following afforestation of former farmland[J]. Forest Ecol Manag,2016,376:59−66. doi: 10.1016/j.foreco.2016.06.004
[33] 宁志英,李玉霖,杨红玲,孙殿超,毕京东. 科尔沁沙地主要植物细根和叶片碳、氮、磷化学计量特征[J]. 植物生态学报,2017,41(10):1069−1080. doi: 10.17521/cjpe.2017.0048 Ning ZY,Li YL,Yang HL,Sun DC,Bi JD. Carbon,nitrogen and phosphorus stoichiometry in leaves and fine roots of dominant plants in Horqin sandy land[J]. Chinese Journal of Plant Ecology,2017,41 (10):1069−1080. doi: 10.17521/cjpe.2017.0048
[34] 胡启武,聂兰琴,郑艳明,吴琴,尧波,郑林. 沙化程度和林龄对湿地松叶片及林下土壤C、N、P化学计量特征影响[J]. 生态学报,2014,34(9):2246−2255. Hu QW,Nie LQ,Zheng YM,Wu Q,Yao B,Zheng L. Effects of desertification intensity and stand age on leaf and soil carbon,nitrogen and phosphorus stoichiometry in Pinus elliottii plantation[J]. Acta Ecologica Sinica,2014,34 (9):2246−2255.
[35] Reed SC,Townsend AR,Davidson EA,Cleveland CC. Stoichiometric patterns in foliar nutrient resorption across multiple scales[J]. New Phytol,2012,196 (1):173−180. doi: 10.1111/j.1469-8137.2012.04249.x
[36] 邱岭军,胡欢甜,林宇,葛露露,王柯远,等. 滨海沙地不同林龄尾巨桉内吸收率及其C∶N∶P化学计量特征[J]. 应用与环境生物学报,2017,23(4):739−744. Qiu LJ,Hu HT,Lin Y,Ge LL,Wang KY,et al. Nutrient resorption efficiency and C∶N∶P stoichiometry of Eucalyptus urophylla × E. grandis of different ages in a sandy coastal plain area[J]. Chinese Journal of Applied and Environmental Biology,2017,23 (4):739−744.
[37] Venterink HO,Wassen MJ,Verkroost AWM,De Ruiter PC. Species richness-productivity patterns differ between N-,P-,and K-limited wetlands[J]. Ecology,2003,84 (8):2191−2199. doi: 10.1890/01-0639
[38] Koerselman W,Meuleman AFM. The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation[J]. J Appl Ecol,1996,33 (6):1441−1450. doi: 10.2307/2404783
[39] Yuan ZY,Chen HYH. Global-scale patterns of nutrient resorption associated with latitude,temperature and PREcipitation[J]. Global Ecol Biogeogr,2009,18 (1):11−18. doi: 10.1111/j.1466-8238.2008.00425.x
[40] 陆姣云,段兵红,杨梅,杨晗,杨惠敏. 植物叶片氮磷养分重吸收规律及其调控机制研究进展[J]. 草业学报,2018,27(4):178−188. Lu JY,Duan BH,Yang M,Yang H,Yang HM. Research progress in nitrogen and phosphorus resorption from senesced leaves and the influence of ontogenetic and environmental factors[J]. Acta Prataculturae Sinica,2018,27 (4):178−188.
[41] Vergutz L,Manzoni S,Porporato A,Novais RF,Jackson RB,Henry H. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants[J]. Ecol Monogr,2012,82 (2):205−220. doi: 10.1890/11-0416.1
[42] 宁秋蕊,李守中,姜良超,陶晶晶,陈涵睿,等. 植物叶片养分再吸收特征及其影响因子[J]. 应用与环境生物学报,2017,23(5):811−817. Ning QR,Li SZ,Jiang LC,Tao JJ,Chen HR,et al. Characteristics and factors influencing foliar nutrient resorption in plants[J]. Chinese Journal of Applied and Environmental Biology,2017,23 (5):811−817.
[43] 郭浩,庄伟伟,李进. 古尔班通古特沙漠中4种荒漠草本植物的生物量与化学计量特征[J]. 植物研究,2019,39(3):421−430. Guo H,Zhuang WW,Li J. Characteristics of biomass and stoichiometry of four desert herbaceous plants in the Gurbantunggut desert[J]. Bulletin of Botanical Research,2019,39 (3):421−430.
[44] 林宇. 厚荚相思N、P内吸收率及其化学计量特征[J]. 福建林业科技,2019,46(1):6−11. Lin Y. Nutrient resorption efficiency and stoichiometry of N and P on Acacia crassicarpa plantations[J]. Journal of Fujian Forestry Science and Technology,2019,46 (1):6−11.
[45] 张燕. 磷添加对青海扁茎早熟禾养分重吸收的影响[D]. 西宁: 青海大学, 2021: 53−55. [46] 魏大平,张健,张丹桔,李川北,赵燕波,等. 不同林冠郁闭度马尾松(Pinus massoniana)叶片养分再吸收率及其化学计量特征[J]. 应用与环境生物学报,2017,23(3):560−569. Wei DP,Zhang J,Zhang DJ,Li CB,Zhao YB,et al. Leaf carbon,nitrogen,and phosphorus resorption and the stoichiometry in Pinus massoniana plantations with various canopy densities[J]. Chinese Journal of Applied and Environmental Biology,2017,23 (3):560−569.
[47] Han WX,Tang LY,Chen YH,Fang JY. Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants[J]. PLoS One,2013,8 (12):e83366. doi: 10.1371/journal.pone.0083366
[48] Liu CC,Liu YG,Guo K,Wang SJ,Yang Y. Concentrations and resorption patterns of 13 nutrients in different plant functional types in the karst region of south-western China[J]. Ann Bot,2014,113 (5):873−885. doi: 10.1093/aob/mcu005
[49] 徐昕,洪江涛,王小丹. 藏北两种牧草添加氮磷后叶片N∶P计量学特征及重吸收率[J]. 草业科学,2022,39(1):85−95. Xu X,Hong JT,Wang XD. Effects of nitrogen and phosphorus addition on leaf N∶P stoichiometry and resorption efficiency of Hordeum vulgare and Elymus nutans in northern Tibet[J]. Pratacultural Science,2022,39 (1):85−95.
[50] Kobe RK,Lepczyk CA,Iyer M. Resorption efficiency decreases with increasing green leaf nutrients in a global data set[J]. Ecology,2005,86 (10):2780−2792. doi: 10.1890/04-1830
[51] Zhou LL,Addo-Danso SD,Wu PF,Li SB,Zou XH,et al. Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry of Cunninghamia lanceolata with stand development in southern China[J]. J Soil Sediments,2016,16 (5):1448−1459. doi: 10.1007/s11368-016-1352-2
[52] Tang LY,Han WX,Chen YH,Fang JY. Resorption proficiency and efficiency of leaf nutrients in woody plants in eastern China[J]. J Plant Ecol,2013,6 (5):408−417. doi: 10.1093/jpe/rtt013
-
期刊类型引用(4)
1. 包兴胜,刘平山,王志明,刘欣,车建美,郑雪芳,王阶平,吴仁烨,刘波. 菌草家园—微生物农业综合生产系统的构建. 中国农学通报. 2025(05): 119-129 .  百度学术
百度学术
2. 陈昱宇,余振东,吴方圆,谢伟东,吕琪,王椿云. 不同油茶品系叶片C、N、P生态化学计量及其重吸收效率研究. 热带农业科学. 2024(02): 19-25 . 百度学术
3. 赵睿,刘玉臣,沈海龙,张鹏,张耀明,高明,郭文慧,闫平. 林木叶片养分重吸收影响因素. 温带林业研究. 2024(04): 89-94 . 百度学术
4. 钟小瑛,任奕炜,衣华鹏,祝贺,高猛. 烟台海岸带地区狗尾草内稳性缓解P限制. 中国环境科学. 2023(10): 5498-5507 . 百度学术
其他类型引用(3)

计量
- 文章访问数: 201
- HTML全文浏览量: 26
- PDF下载量: 44
- 被引次数: 7
