
Characteristics of Phyllostachys edulis (Carrière) J. Houz shoot morphology, branching, and leaf expansion efficiency during understory vegetation succession
-
摘要:
本研究分别选取林下植被演替前毛竹(Phyllostachys edulis (Carrière) J. Houz)纯林经营、竹材6~8年采伐一次形成的林下植被演替9、21年以及纯林(对照)等3类毛竹林,调查分析了1度、2度竹的主侧枝枝叶形态质量、抽枝展叶效率及生物量异速生长关系等。结果显示:(1)随林下植被演替的进行,1度竹主枝叶片主要形态质量性状呈降低趋势,2度竹则呈“V”型变化;(2)1度、2度竹主侧枝的枝长、纤细率显著提高,枝基径、枝干物质含量、枝干重等性状呈倒“V”型或升高趋势,枝形态变化较枝质量变化明显;(3)1度竹抽枝展叶效率指标呈“V”型变化,而2度竹主侧枝抽枝展叶效率显著降低,胡波尔值明显提高;(4)一定演替年限内,立竹主侧枝枝叶生物量生长关系从等速生长向异速生长变化,尔后恢复为等速生长。可见,林下植被演替对毛竹林立竹枝叶形态质量、抽枝展叶效率和枝叶生物量异速生长关系均产生重要影响,叶片趋于变小,枝条趋于细长,且具有明显的演替年限效应和立竹年龄效应,主要对主枝叶片产生影响,1度竹较2度竹敏感;演替过程中立竹将更多的资源投资于枝的生长,以增强空间资源的竞争能力。
Abstract:This research examined three types of moso bamboo stands: pure moso bamboo forest prior to understory vegetation succession, understory vegetation succession formed by harvesting bamboo timber once every 6–8 years over periods of 9 and 21 years, and pure forest (control). We investigated the morphological quality of the main and lateral branches and leaves, efficiency of pumping and spreading, and biomass anisotropic growth relationship of 1st- and 2nd-degree bamboo. Results showed that: (1) With the succession of understory vegetation, the primary morphological and quality traits of the main branch leaves of 1st-degree bamboo showed a downward trend, while 2nd-degree bamboo showed a significant “V”-shaped change; (2) Length and slenderness of the main and lateral branches of 1st- and 2nd-degree bamboo increased significantly, while branch diameter, branch dry matter content, and branch dry weight showed an inverted “V”-shaped or increasing trend, with the changes in branch morphology being more pronounced than those in branch quality; (3) The leaf spreading efficiency index of 1st-degree bamboo showed a “V”-shaped change, whereas that of the main and lateral branches of 2nd-degree bamboo decreased significantly, with the Huber value increasing significantly; (4) Within a certain succession period, the biomass growth relationship of the main and lateral branches of the standing bamboo transitioned from isochronous to anisotropic growth, then reverted to isochronous growth. Thus, understory vegetation succession exhibited an important effect on the morphological quality of bamboo branches and leaves, efficiency of branching and spreading, and relationship between anisotropic growth and branch and leaf biomass in moso bamboo forests. Notably, leaves tended to be smaller and branches slenderer. Furthermore, successional age and age of standing bamboo had obvious effects, mainly on the leaves of the main branches, with 1st-degree bamboo being more sensitive than 2nd-degree bamboo. Hence, during the succession process, bamboo invests more resources in branch growth to enhance competition for spatial resources.
-
土壤重金属污染是各方关注的严重环境问题之一。根据2014年《全国土壤重金属污染状况调查公报》,我国约有2 000万公顷耕地受到了重金属污染,其中330万公顷的污染程度达到了中度或重度污染。江西是我国矿产资源大省,铜资源在全国具有重要地位,长期的矿业开采、选矿、冶炼等产生的“三废”不断排放到周围环境,造成周边农田土壤铜污染严重,对农产品安全和人类身体健康都造成了极大的威胁[1]。以江西德兴铜矿区为例,周边土壤存在不同程度的铜污染,在919份调查样品中,铜含量位于50~400 mg/kg的有171个,含量大于400 mg/kg有18个,土壤最高铜含量可达1825 mg/kg,远大于中国土壤环境质量标准的三级标准(400 mg/kg)[2, 3],铜污染土壤的治理逐渐成为一个亟待解决的问题。植物修复(Phytoremediation)是一种新兴的土壤重金属净化技术[4],具有成本低廉、修复面积广、操作方便、二次污染易控制,美化植被环境等优点[5]。目前,关于修复植物的筛选多集中在草本植物,但草本生物量小、生长速度慢且很难进行机械收获作业,不适宜开展大面积污染土壤的修复[6],且农作物和草类等所吸收的重金属大多存在进入食物链或二次污染的风险,因此在实际修复应用中受到限制。当前,寻找生物量大、适应性强、观赏价值高且耐重金属的植物十分具有现实意义[7,8]。木本植物对重金属具有较强的耐性和一定的固定、吸收、累积能力,且其发达的深根系可使土壤中重金属移动性降低,降低其迁移和生物毒性,改善土壤质量等,引起研究人员的广泛关注[9]。
樟树(Cinnamomum camphora (L.) Presl)是重要的木本经济树种,樟脑、樟油等提取物是医药、化工、香料、防腐、农药的重要原料。樟树具有根系发达、生长迅速、生物量大、适应性强等优点,对重金属污染环境有较强的适应能力,15种常见乔木中,樟树对重金属的综合富集系数最高,对土壤中铜、锌和镉的富集能力均较强[10],在铜为主要污染源的土壤植物修复中有着较大的应用潜力。因此,樟树具有土壤修复收益型植物潜力。项目组前期将全国范围内收集的9个种源68个家系的樟树实生苗种植在以铜污染为主的江西德兴铜矿周边的水龙山实验修复基地,经过3年的跟踪调查,有12个家系适应性表现良好,成活率和保存率均较高,说明部分樟树具有较好的耐铜性,但樟树对铜离子的生理生化响应及富集转运特性还不清楚。本项目以前期筛选出的耐铜樟树为实验材料,通过盆栽实验研究不同浓度铜处理对樟树生长和生理指标的影响及铜的富集转运特性,探明耐铜樟树抵御铜毒性的生理生化机制,以期为樟树用于铜污染土壤修复提供科学依据。
1. 材料与方法
1.1 实验材料
实验于2021年7月在江西省科学院生物资源研究所人工气候室进行。实验地位于南昌市(28°41'N,115°59'E),实验材料为前期筛选出的耐铜樟树材料。收集同一单株的种子,将种子去果肉并反复搓洗干净,室内沙藏,于第2年春天播种,筛选长势一致的樟树幼苗统一移栽至事先准备好的基质中(红壤土∶细沙∶泥炭=1∶1∶1),栽培盆大小为上口径20 cm,底径14 cm,高14 cm,每盆基质重2.5 kg。经过3个月的缓苗,选取生长较好、高度一致的幼苗,采用随机区组设计,进行铜胁迫处理。
1.2 实验方法
以CuSO4·5H2O 为铜来源,配制铜浓度为100 g/kg的母液,处理前1周各栽培盆开始控水,至处理时各盆中基质均为干燥状态,处理时将各处理浓度所需母液用蒸馏水稀释至550 mL,缓缓浇灌至栽培盆中(经实验前反复验证,550 mL可以保证栽培盆内基质完全浸湿,且托盘里几乎没有多余水分流出),处理浓度分别为:0、50、150、300、600、900、1200 mg/kg(以基质重计算纯铜含量),每个处理4个重复。采样时间为处理60 d,每株选取3~4片嫩叶(枝条顶部开始倒数第 3~4 片叶)为待测材料,以锡箔纸包好,迅速投入液氮罐中,带回实验室放入−80 ℃冰箱保存,用于生理指标的测定;另准确称取0.1 g新鲜叶片,剪碎,放入事先准备好的装有95%乙醇的离心管中,黑暗处理,并放在摇床上震荡处理3 d,至叶片完全透明为止,用于色素含量的测定;最后将各处理单株分地上地下保存,放烘箱105 ℃杀青30 min,再70 ℃烘干至恒重测定干物质量,每株分别准确称取0.2 g根、茎、叶的研磨粉,微波消解,采用ICP-MS测定不同器官中铜含量。
1.3 生理指标测定
生理指标检测均参考施海涛[11]的方法。氯化硝基四氮唑蓝(NBT)光还原法测超氧化物歧化酶(SOD)活性;愈创木酚氧化法测过氧化物酶(POD)活性;高锰酸钾滴定法测过氧化氢酶(CAT)活性;烟酰胺腺嘌呤二核苷酸磷酸还原反应法测谷胱甘肽还原酶(GR)活性;硫代巴比妥酸比色法测丙二醛(MDA)含量;茚三酮比色法间接测定游离脯氨酸(Pro))含量;蒽酮比色法测定可溶性糖含量;考马斯亮蓝法测定可溶性蛋白含量。硫酸钛反应法测定过氧化氢(H2O2)含量,2-硝基苯甲酸反应法测谷胱甘肽(GSH)含量。所用试剂盒均由南京建成生物工程研究所提供,具体操作步骤按照相关说明书进行。
1.4 数据处理
所有指标的测定均进行4次生物学重复,最后结果采用平均值±标准差表示,采用SPSS 25软件作图并进行单因素方差分析(One-way ANOVA)。
2. 结果与分析
2.1 铜胁迫对樟树生长与生物量积累的影响
如图1所示,在铜胁迫浓度小于600 mg/kg时,樟树都可以很好地存活,根系和叶片均没有明显变化。其中150 mg/kg铜处理时,樟树长势最好,茎秆粗壮,叶片颜色墨绿。铜胁迫浓度为900 mg/kg时,叶片没有明显变化,部分根系开始出现褐化,根系的数量开始减少,但处理2个月后有新根萌出,可见900 mg/kg处理时,樟树依然可以存活。当处理浓度为1 200 mg/kg时,部分叶片开始萎蔫,根系褐化严重,且根系数量减少明显,也没有新根长出。因此在1 200 mg/kg铜处理下,樟树生长抑制及损害明显。
![]() 图 1 不同浓度铜处理60 d时樟树形态的变化1~7分别为0、50、150、300、600、900、1 200 mg/kg铜浓度处理。Figure 1. Morphological changes in Cinnamomum camphora treated with different concentrations of copper for 60 d1–7 were treated with 0, 50, 150, 300, 600, 900, and 1 200 mg/kg copper concentrations, respectively.
图 1 不同浓度铜处理60 d时樟树形态的变化1~7分别为0、50、150、300、600、900、1 200 mg/kg铜浓度处理。Figure 1. Morphological changes in Cinnamomum camphora treated with different concentrations of copper for 60 d1–7 were treated with 0, 50, 150, 300, 600, 900, and 1 200 mg/kg copper concentrations, respectively.生物量干质量的结果与生长观察一致(表1),150 mg/kg处理时可以促进樟树的生长,其地上和根系干质量均高于其他处理,900 mg/kg处理时,地上干质量没有降低,反而高于对照组,但差异不显著(P>0.05);根系干质量稍有降低,差异也不显著(P>0.05)。1 200 mg/kg处理时,樟树的地上和根系干质量均低于其他各处理,且与150 mg/kg处理时差异显著(P<0.05),与其他处理差异不显著(P>0.05)。
表 1 不同浓度铜处理下樟树地上和根系生物量(干质量)的测定Table 1. Determination of aboveground and root biomass (dry mass) of Cinnamomum camphora treated with different concentrations of copper处理
Treatments / mg/kg地上质量
Dry mass of shoot / g根质量
Dry mass of root / g0 7.98±2.87bc 3.81±1.18ab 50 7.85±2.54bc 3.59±0.67ab 150 9.59±3.08a 4.53±1.35a 300 9.18±1.57ab 3.62±0.53ab 600 7.61±1.14bc 3.49±0.50ab 900 8.33±0.91bc 3.11±0.60b 1200 6.82±1.44c 2.97±0.90b 注:数据均为平均值±标准差,小写字母表示不同处理间在P<0.05水平上差异显著(n=4)。下同。 Notes: All data in the table are average values±standard deviation (SD). Lowercase letters after data in the same row indicate significant differences between different treatments at P<0.05 level (n=4). Same below. 2.2 铜胁迫对樟树光合色素含量的影响
研究结果显示,叶绿素a、b及类胡萝卜素的含量均随着铜浓度的升高而降低。处理浓度为1200 mg/kg时,叶绿素a、b的含量均降至最低,显著低于其他各处理(P<0.05,图2);900 mg/kg处理时,叶绿素a、b的含量与300、600 mg/kg处理的差异均不显著(P>0.05),但与CK、50、150 mg/kg 处理时差异显著(P<0.05)。50、150、300、600 mg/kg 4种处理间的叶绿素a与叶绿素b差异均不显著(P>0.05)。类胡萝卜素含量的变化趋势与叶绿素a、b的变化趋势大致相同,也表现出随处理浓度的升高而降低的趋势,1 200 mg/kg处理时,类胡萝卜素含量最低,但与50、600、900 mg/kg 3个处理的差异不显著(P>0.05),与CK、150、300 mg/kg处理的差异显著(P<0.05)。
![]() 图 2 不同浓度铜处理60 d叶片中叶绿素a、b及类胡萝卜素含量的测定不同小写字母表示处理间在 P<0.05水平差异显著。下同。Figure 2. Determination of chlorophyll a, chlorophyll b, and carotenoid in Cinnamomum camphora leaves treated with different concentrations of copper for 60 dDifferent lowercase letters indicate significant differences between different treatments at P<0.05 level. Same below.
图 2 不同浓度铜处理60 d叶片中叶绿素a、b及类胡萝卜素含量的测定不同小写字母表示处理间在 P<0.05水平差异显著。下同。Figure 2. Determination of chlorophyll a, chlorophyll b, and carotenoid in Cinnamomum camphora leaves treated with different concentrations of copper for 60 dDifferent lowercase letters indicate significant differences between different treatments at P<0.05 level. Same below.2.3 铜胁迫对樟树抗氧化酶系统的影响
不同铜浓度影响樟树抗氧化酶系统的分析结果表明(图3),CAT活性在150 mg/kg时最低,与CK、50、600、900、1 200 mg/kg处理的差异显著(P<0.05;图3:A), CK和1 200 mg/kg两个处理时CAT活性最高,但与50、600、900 mg/kg处理间的差异不显著(P>0.05)。POD活性在CK、150、900 mg/kg处理时最高,显著高于300和600 mg/kg两个处理,600 mg/kg处理时的POD活性最低(图3:B)。SOD活性最高的是CK和150 mg/kg两个处理,显著高于300、600 mg/kg处理,与其他处理间的差异不显著(图3:C)。谷胱甘肽还原酶(GR)活性有随着处理浓度的升高逐渐降低的趋势,CK和50 mg/kg处理的GR活性最高,1 200 mg/kg处理时,活性最低,与CK、50、150、300、600 mg/kg处理差异显著,但与900 mg/kg处理的差异不显著(图3:D)
![]() 图 3 不同浓度铜处理60 d樟树叶的酶活指标变化Figure 3. Changes in enzyme activity indices of Cinnamomum camphora leaves treated with different concentrations of copper for 60 d
图 3 不同浓度铜处理60 d樟树叶的酶活指标变化Figure 3. Changes in enzyme activity indices of Cinnamomum camphora leaves treated with different concentrations of copper for 60 d2.4 铜胁迫对樟树渗透调节物质含量及抗逆性指标的影响
GSH作为植物体内重要的抗氧化剂和自由基清除剂,有随着处理浓度的升高而逐渐升高的趋势(图4:A)。渗透调节物质脯氨酸和可溶性糖含量均在1 200 mg/kg 处理时显著高于其他处理(P<0.05),其他处理间没有显著差异(P>0.05;图4:B、C)。可溶性蛋白在900和1 200 mg/kg 处理时显著高于其他处理,其他处理间差异不显著(图4:D)。随着铜浓度的升高,抗逆性指标MDA表现出逐渐升高的趋势(图4:E);H2O2浓度在CK和1 200 mg/kg两个处理时达到最高水平,显著高于50、150、300、600 mg/kg处理,与900 mg/kg处理的差异不显著(图4:F)。
![]() 图 4 不同铜浓度处理60 d时樟树叶的抗逆性生理指标变化Figure 4. Changes in stress resistance physiological indices of Cinnamomum camphora leaves treated with different concentrations of copper for 60 d
图 4 不同铜浓度处理60 d时樟树叶的抗逆性生理指标变化Figure 4. Changes in stress resistance physiological indices of Cinnamomum camphora leaves treated with different concentrations of copper for 60 d2.5 铜胁迫对樟树不同组织铜浓度的影响
由表2可知,随着铜浓度的增加,根、茎、叶等不同组织的铜含量也逐渐升高,但与CK相比,处理浓度为50和150 mg/kg时,差异没有达到显著水平(P>0.05)。当处理浓度在300 mg/kg以上时,樟树叶、茎和根的铜含量均显著高于对照,但叶、茎的增幅显著低于根,与对照处理相比,300、600、900、1 200 mg/kg处理的叶片铜含量分别增加了1.73、1.80、1.74和2.12倍,茎部铜含量分别增加了1.42、1.45、1.35和2.02倍,而根部铜含量则分别增加了4.45、3.95、15.72和22.17倍。随着铜浓度的增加,铜离子由根系向叶片的转移率IF(Leaf/Root)由对照处理的0.078逐渐降至1 200 mg/kg处理时的0.007;铜离子由根系向茎的转移率IF(Stem/Root)由对照处理的0.06逐渐降至1 200 mg/kg处理时的0.005。樟树各器官铜含量的大小顺序为根>叶>茎,樟树主要将铜富集在根系,往地上茎和叶的转移率很低,且随着铜浓度的增大,往地上茎和叶的转移率也显著降低。
表 2 不同铜浓度处理下樟树不同部位铜含量及铜转移率的测定Table 2. Effects of different concentrations of copper on copper content and transfer rate in different parts of Cinnamomum camphora处理
Treatments /
mg/kg叶铜含量
Leaf Cu /
mg/kg茎铜含量
Stem Cu /
mg/kg根铜含量
Root Cu /
mg/kg铜转运
(叶/根)
IF (Leaf/Root)铜转运
(茎/根)
IF (Stem/Root)0 1.91±0.13c 1.46±0.29c 24.46±1.34d 0.078±0.005a 0.060±0.011a 50 1.72±0.01c 1.09±0.13c 25.20±1.28d 0.068±0.004b 0.044±0.007b 150 1.96±0.45c 1.40±0.23c 28.80±3.16d 0.067±0.009b 0.049±0.011ab 300 3.32±0.43b 2.08±0.24b 108.95±13.01c 0.031±0.002c 0.019±0.003c 600 3.44±0.37b 2.12±0.41b 96.67±11.71c 0.036±0.008c 0.022±0.006c 900 3.33±0.66b 1.97±0.36b 384.44±34.11b 0.009±0.002d 0.005±0.001d 1 200 4.06±0.53a 2.95±0.01a 542.27±51.01a 0.007±0.001d 0.005±0.001d 3. 讨论
3.1 铜胁迫对樟树生长的影响
樟树是重要的木本经济树种,茎叶是精油提取的主要器官,若能将樟树用于铜污染土壤修复中,不仅能改善生态环境,还可以实现收益,同时具有经济和生态效益。研究发现,樟树对铜污染有较好的适应能力[12, 13],在铜污染土壤的植物修复中应用潜力较大[14, 15]。本研究中,樟树对铜胁迫表现出“低促高抑”的变化趋势,一定浓度的铜(150 mg/kg)促进樟树的生长,其地上和根系生物量均高于其他处理,900 mg/kg处理时,樟树地上干质量没有降低,根系干重稍有降低,当处理浓度为1 200 mg/kg时,樟树的地上和根系干质量均降低。生长状况是反映植物对重金属耐受程度的直观指标[7],过量铜抑制植物生长曾多次被文献报道,例如,当Cu2+浓度高于20 μmol/L时,小白菜(Brassica campestris L. ssp. chinensis Makino)和芥菜(Brassica juncea L. Czern. et Coss.)的根、茎、叶生物量均随着铜浓度的升高而显著降低,铜胁迫抑制了小白菜和芥菜的生长[16, 17];玉米(Zea mays L.)幼苗的铜胁迫处理表明,Cu2+浓度为5 μmol/L时玉米幼苗的生长即受到抑制,随着胁迫浓度的升高,玉米幼苗地上和地下生物量均显著降低[18]。
光合作用是植物获取能量的重要途径,植物的能量含量与光合作用密切相关,叶绿素含量是反映植物耐受逆境胁迫的重要指标之一[19],本研究中叶绿素a、b及类胡萝卜素的含量均有随着铜处理浓度的升高而降低的趋势,与袁霞等[20]、李炎林等[21]的研究结果一致。当铜浓度为900 mg/kg时,叶绿素含量与其他低浓度处理差异均不显著;当铜浓度为1 200 mg/kg时,叶绿素含量则显著低于其他各处理。原因可能是重金属胁迫下,植物叶绿体膜结构受到一定程度的破坏,叶绿体开始降解,且叶绿体内的光合酶合成受抑制,叶片气孔关闭,光合作用强度降低[22],植物的生物量积累减少,生长受到抑制。
3.2 铜胁迫对樟树生理生化反应的影响
铜胁迫导致叶绿素含量降低及生长受抑制的主要原因是细胞内活性氧(ROS)含量不断增加并积累[23, 24]。低浓度的重金属可以激活植物的抗氧化应急系统,诱导 SOD、POD和CAT活性升高,以清除体内的活性氧[25],高浓度的 ROS则破坏细胞膜、核酸和叶绿素等关键成分,造成氧化损伤,影响植物的正常发育[26, 27]。MDA是细胞膜脂过氧化的产物,含量的高低反映了植物细胞膜脂过氧化的程度以及抗逆能力的强弱[25],MDA含量越高,植物细胞膜脂过氧化程度越高,ROS引起的细胞损伤越大[28]。本研究中MDA含量有随着铜处理浓度的升高而升高的趋势,900和1 200 mg/kg两个浓度处理时,MDA的含量显著高于其他各处理,说明高浓度铜胁迫对樟树细胞造成一定程度损伤。
可溶性蛋白、可溶性糖和游离脯氨酸作为植物体内重要的渗透调节物质,其含量的变化也是植物抗逆性强弱的重要反映。可溶性蛋白是植物细胞中含量最丰富的生物大分子之一,是植物结构和功能的重要物质基础,含量越高说明生理生化反应和代谢活动越强[29, 30]。本研究中,可溶性蛋白在铜浓度小于600 mg/kg时含量相对稳定,在900和1 200 mg/kg两个高浓度处理时,蛋白含量显著高于其他各处理,说明900和1 200 mg/kg两个浓度处理时,樟树的生理生化和代谢活动显著增强。游离脯氨酸既是渗透调节剂,又可作为自由基清除体内的重金属,其含量的增加可提高植物细胞液的浓度,维持细胞内外渗透压的平衡,保障细胞膜的结构稳定[31]。可溶性糖在维持细胞膜稳定、细胞扩张、酶活性和植株光合作用等方面起着重要作用[32],植物在逆境中可以通过增加可溶性糖等的含量来调节细胞的渗透压,提高对逆境的适应能力[33]。本研究中游离脯氨酸和可溶性糖的含量均在1 200 mg/kg处理时达到最高水平,显著高于其他各处理,900 mg/kg处理和其他低浓度处理的差异不显著,说明900 mg/kg处理时,细胞的内环境相对稳定。
植物抵抗重金属的能力与体内抗氧化酶系统的活性密切相关。POD、 CAT 与 SOD 协同作用,能够有效清除胁迫产生的活性氧,减少对植物细胞的危害,保证植物的正常生长[34]。过氧化氢是在铜胁迫下产生的活性氧物质,能直接或间接氧化细胞内的核酸、蛋白质等生物大分子,破坏细胞膜结构,从而加速细胞老化和解体,POD通过催化过氧化氢分解成无毒害的水和氧气,使植物体内的自由基保持在较低水平,从而避免对植物造成氧化伤害[35,36]。本研究中,不同铜处理后SOD、POD 和 CAT 酶的活性没有显著升高,过氧化氢的含量也相对稳定,在900 mg/kg处理时,其含量与对照差异不显著,说明该处理浓度没有超出樟树的自身调节能力,樟树可以有效抵抗铜的毒害作用。
3.3 铜胁迫对樟树不同器官铜含量的影响
植物对重金属的富集和转运能力是评价其耐受性的重要指标,与根系吸收及根系向地上部转运能力密切相关[9, 37]。本研究中,樟树主要将铜富集在根系,往地上茎和叶的转移率很低,且随着铜浓度的增加,往地上茎和叶的转移率逐渐降低,减轻了过量铜对樟树地上组织的毒害作用,与朱宇恩等[38]的研究结果一致。研究发现,植物主要通过两种途径抵抗铜的毒害作用:一是减少铜从根向地上部的转移,铜离子进入根系后,更多的集中在根系,从而降低铜对植物地上部的毒性;二是将铜离子集中在对铜毒性不太敏感的组织或部位固定起来,以减少铜在植物细胞中的转移,主要方式有将铜离子结合在细胞壁上或者转移到液泡中[39],或者利用植物体内的螯合剂与铜离子结合,形成对植物毒性较小或无毒的螯合物[40]。GSH为植物螯合剂(PCs)的合成提供合成前体,GR能够还原氧化型谷胱甘肽(GSSG)生成还原型谷胱甘肽(GSH),维持细胞内充足的GSH水平[41]。 本研究中,GSH含量有随着铜浓度的升高而升高的趋势,在900和1 200 mg/kg两个高浓度处理时,显著高于其他各处理;GR活性则在CK和50 mg/kg处理时活性最高,且随着处理浓度的升高逐渐降低,1 200 mg/kg处理时活性最低,推测GR被大量消耗,以合成更多的GSH参与植物螯合剂的合成并螯合过量铜离子。 但目前为止,樟树根系对铜离子的耐受策略及其分子调控机制还不清楚,深入探究樟树根系对铜离子的耐受和转运机制,是我们下一步要重点研究的内容。
-
![]()
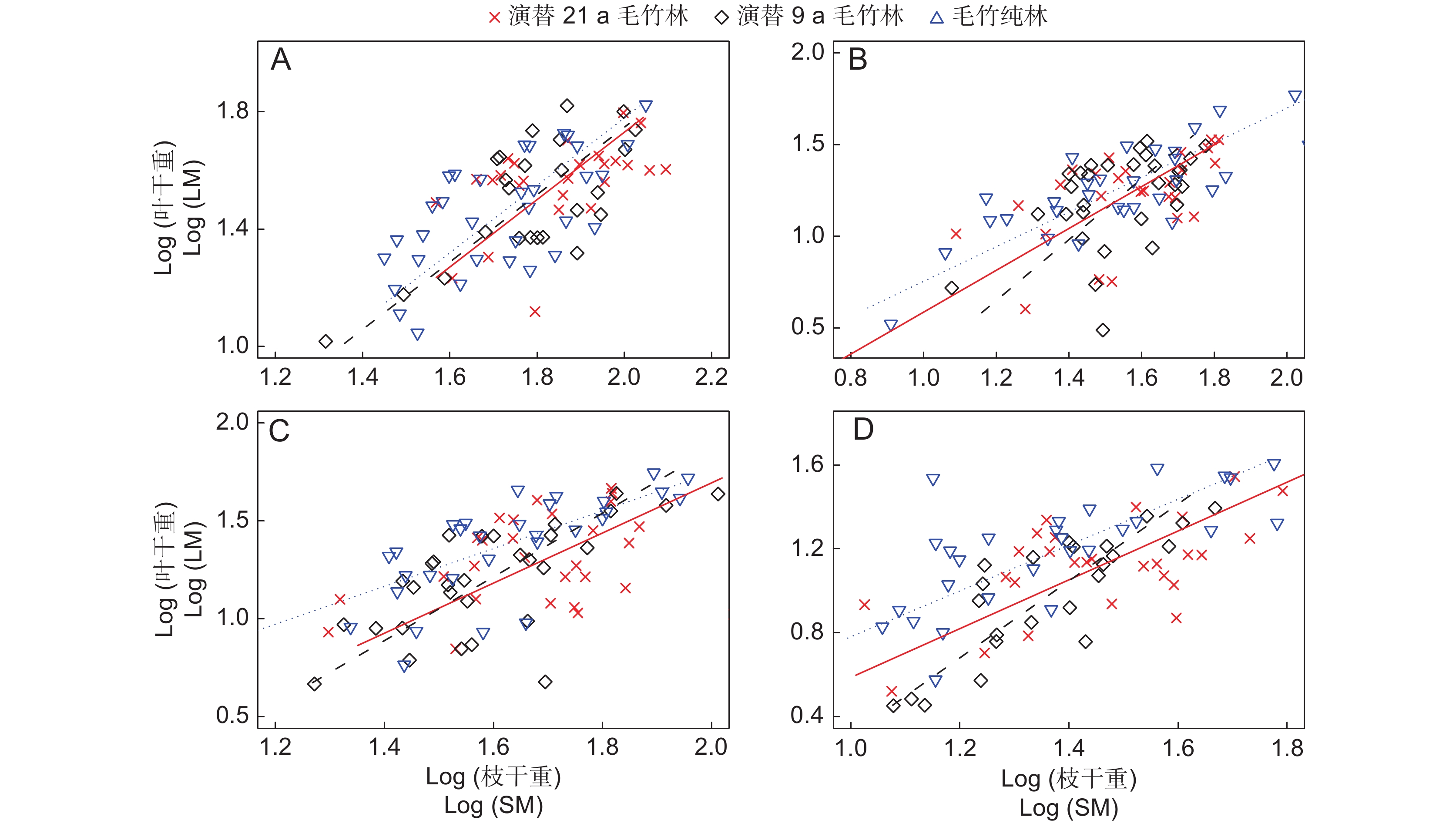
图 1 毛竹林叶干重与枝干重异速生长关系
A:1度竹主枝;B:1度竹侧枝;C:2度竹主枝;D:2度竹侧枝。
Figure 1. Relationship between total leaf mass and stem mass of experimental Phyllostachys edulis stands
A: Main branches of 1st-degree bamboo; B: Lateral branches of 1st-degree bamboo; C: Main branches of 2nd-degree bamboo; D: Lateral branches of 2nd-degree bamboo.
表 1 实验毛竹林基本情况
Table 1 Basic information on experimental Phyllostachys edulis stands
演替年限
Succession
period / a海拔
Altitude / m坡向
Slope
aspect坡位
Slope
position毛竹
P. edulis乔灌木
Arbor and shrubs郁闭度
Canopy
density立竹密度
Bamboo
density /
ind/hm2立竹胸径
Bamboo diameter
at breast height / cm密度
Density /
ind/hm2树高
Tree height / m地径
Basal
diameter / cm0 500 ± 10 36 ± 2.50 西 2 625 ± 25 9.40 ± 0.60 - - - 0.60 9 520 ± 10 36 ± 2.50 西 3 150 ± 30 9.20 ± 0.40 8 000 ± 55 0.76 ± 0.08 0.76 ± 0.07 0.70 21 490 ± 10 36 ± 2.50 西 2 275 ± 20 8.90 ± 0.40 27 511 ± 147 2.42 ± 0.20 1.69 ± 0.11 0.85  下载: 导出CSV
下载: 导出CSV
表 2 毛竹林立竹叶片形态质量性状
Table 2 Leaf morphological characteristics of experimental Phyllostachys edulis stands
演替年限
Succession
period / a立竹年龄
Bamboo
age枝类
Branch
class叶长
Leaf
length / cm叶宽
Leaf
width / cm叶周长
Leaf
circumference / cm叶干重
Leaf dry
weight / g单叶面积
Leaf
area / cm2比叶面积
Specific leaf
area / cm2/g叶干物质含量
Leaf dry
matter content /
g/g21 1 主枝 9.63 ± 0.28B 1.44 ± 0.03B 22.02 ± 0.30B 33.94 ± 1.79A 10.31 ± 0.30B 195.53 ± 2.61A 0.12 ± 0.00A 9 9.88 ± 0.45B 1.45 ± 0.03B 22.27 ± 0.91B 34.23 ± 1.08A 10.60 ± 0.67B 174.21 ± 1.73B 0.14 ± 0.01A 0 10.67 ± 0.31A 1.52 ± 0.03A 24.18 ± 0.48A 30.51 ± 0.07B 11.97 ± 0.34A 200.31 ± 5.13A 0.13 ± 0.01A 21 侧枝 9.94 ± 0.21A 1.46 ± 0.03A 22.65 ± 0.45A 16.93 ± 0.68A 10.81 ± 0.24A 200.03 ± 11.11A 0.12 ± 0.00A 9 10.03 ± 0.26A 1.47 ± 0.03A 22.54 ± 0.69A 18.55 ± 0.16A 10.78 ± 0.39A 169.83 ± 5.15B 0.13 ± 0.01A 0 10.20 ± 0.20A 1.47 ± 0.04A 23.18 ± 0.39A 19.52 ± 2.66A 11.20 ± 0.40A 206.76 ± 8.14A 0.12 ± 0.01A 21 2 主枝 10.66 ± 0.08A 1.54 ± 0.03A 24.26 ± 0.15A 24.37 ± 1.94A 12.34 ± 0.09A 190.20 ± 0.13A 0.15 ± 0.01A 9 9.96 ± 0.12B 1.43 ± 0.04B 22.31 ± 0.02B 19.09 ± 0.74B 10.66 ± 0.23B 182.36 ± 1.95A 0.13 ± 0.01B 0 10.79 ± 0.17A 1.56 ± 0.06A 24.24 ± 0.38 A 27.18 ± 2.00A 12.15 ± 0.19A 191.78 ± 16.56A 0.13 ± 0.00A 21 侧枝 10.56 ± 0.13A 1.49 ± 0.02A 23.41 ± 0.51AB 16.18 ± 1.40B 11.71 ± 0.19A 188.60 ± 0.13A 0.14 ± 0.00A 9 10.02 ± 0.27B 1.47 ± 0.00A 22.58 ± 0.56B 11.02 ± 0.51C 10.98 ± 0.41A 184.19 ± 4.80A 0.15 ± 0.03A 0 10.71 ± 0.20A 1.48 ± 0.02A 24.20 ± 0.53A 19.08 ± 0.89A 11.67 ± 0.48A 190.47 ± 4.64A 0.14 ± 0.00A 注:不同大写字母表示不同林下植被演替年限毛竹林同一年龄立竹主枝或侧枝叶片间差异显著。下同。 Note: Different capital letters indicate different vegetation succession years, with significant differences between the leaves of main and side branches of P. edulis stands of the same age. Same below.
下载: 导出CSV
表 3 毛竹林立竹枝形态质量性状
Table 3 Branch morphological characteristics of experimental Phyllostachys edulis stands
演替年限
Succession
period / a立竹年龄
Bamboo
age枝类
Branch
class枝长
Branch
length / cm枝基径
Branch base
diameter / mm枝干重
Branch
weight / g枝纤细率
Branch fineness
rate / cm/mm枝干物质含量
Branch dry matter
content / g/g21 1 主枝 159.78 ± 10.10A 9.09 ± 0.05A 75.00 ± 1.96A 176.12 ± 6.24A 0.64 ± 0.02A 9 157.22 ± 2.98A 9.18 ± 0.18A 60.20 ± 3.35B 176.05 ± 11.18A 0.64 ± 0.01A 0 143.25 ± 3.47B 8.82 ± 0.27A 55.31 ± 2.25B 163.74 ± 1.19B 0.62 ± 0.02A 21 侧枝 132.67 ± 5.39A 7.12 ± 0.23A 36.82 ± 4.36A 187.16 ± 2.95A 0.64 ± 0.02B 9 128.94 ± 1.59A 6.94 ± 0.07A 36.56 ± 0.52A 189.03 ± 10.30A 0.69 ± 0.05A 0 123.48 ± 6.46B 7.16 ± 0.47A 37.70 ± 3.81A 178.30 ± 6.70B 0.63 ± 0.01B 21 2 主枝 155.42 ± 6.61A 8.23 ± 0.14A 57.75 ± 8.71A 189.53 ± 7.49A 0.61 ± 0.02A 9 137.39 ± 1.42B 7.70 ± 0.14B 43.20 ± 3.95B 180.42 ± 0.70A 0.66 ± 0.01A 0 139.22 ± 1.13B 7.75 ± 0.07B 46.49 ± 3.31B 178.86 ± 5.05B 0.64 ± 0.01A 21 侧枝 137.50 ± 6.50A 6.74 ± 0.19A 32.52 ± 3.55A 203.41 ± 1.92A 0.64 ± 0.01A 9 124.11 ± 2.96B 6.03 ± 0.07B 23.63 ± 0.55B 204.49 ± 4.77A 0.66 ± 0.01A 0 121.31 ± 10.73B 6.59 ± 0.58A 29.60 ± 3.56A 183.98 ± 4.02B 0.64 ± 0.03A
下载: 导出CSV
表 4 毛竹林抽枝展叶效率
Table 4 Branch and leaf characters of experimental Phyllostachys edulis stands
演替年限
Succession
period / a立竹年龄
Bamboo
age枝类
Branch
class出叶强度
Intensity of
leaf / ind/g叶面积比
Leaf area
ratio / mm2/g叶密度
Leaf density /
ind/mm叶枝质量比
Leaf /stem mass
ratio / g/g胡波尔值
Huber value /
mm2/mm21 1 主枝 6.35 ± 0.24A 100.12 ± 5.72B 4.34 ± 0.54A 0.53 ± 0.03B 0.00008 ± 0B 9 5.65 ± 0.56B 93.27 ± 14.77B 3.59 ± 0.76B 0.55 ± 0.08AB 0.00012 ± 0A 0 6.12 ± 0.43AB 117.82 ± 29.07A 3.87 ± 0.70AB 0.61 ± 0.07A 0.00009 ± 0AB 21 侧枝 6.04 ± 0.44B 99.34 ± 10.89B 2.58 ± 0.34A 0.52 ± 0.06B 0.00011 ± 0B 9 5.17 ± 0.28C 84.42 ± 6.93C 2.24 ± 0.30A 0.51 ± 0.04B 0.00012 ± 0A 0 6.68 ± 0.84A 121.27 ± 23.79A 2.73 ± 1.07A 0.61 ± 0.12A 0.00011 ± 0A 21 2 主枝 4.66 ± 0.34C 83.18 ± 8.66B 2.25 ± 0.32B 0.44 ± 0.05B 0.00012 ± 0A 9 5.30 ± 0.44B 84.18 ± 10.19B 2.54 ± 0.53B 0.49 ± 0.06B 0.00013 ± 0A 0 6.16 ± 0.47A 123.26 ± 15.38A 3.49 ± 0.76A 0.64 ± 0.08A 0.00009 ± 0B 21 侧枝 4.97 ± 0.30B 85.52 ± 7.59B 1.68 ± 0.49B 0.47 ± 0.04B 0.00013 ± 0A 9 4.94 ± 0.58B 82.79 ± 14.92 B 1.51 ± 0.27B 0.52 ± 0.09B 0.00014 ± 0A 0 6.72 ± 0.45A 136.35 ± 15.92A 2.67 ± 0.90A 0.73 ± 0.09A 0.00009 ± 0B
下载: 导出CSV
表 5 毛竹林枝-叶质量的SMA分析
Table 5 SMA analysis of branch leaf size of experimental Phyllostachys edulis stands
指标
Indicators演替年限
Succession
period / a立竹年龄
Bamboo
age枝类
Branch
class数量
Number斜率
Slope95%置信区间
95% CI截距
InterceptR2 P 叶干重-
枝干重21 1 主枝 30 1.02(1.14) 0.71,1.45 −0.32 0.25 < 0.010 9 30 1.26(1.14) 0.92,1.72 −0.74 0.48 < 0.001 0 30 1.15(1.14) 0.87,1.51 −0.51 0.42 < 0.001 21 侧枝 30 1.14(1.02) 0.85,1.51 −0.55 0.51 < 0.001 9 30 1.66 1.18,2.32 −1.34 0.23 < 0.050 0 30 0.94(1.02) 0.74,1.20 −0.19 0.61 < 0.001 21 2 主枝 30 1.28(0.99) 0.91,1.81 −0.87 0.36 < 0.050 9 30 1.62 1.23,2.15 −1.39 0.46 < 0.001 0 30 0.97(0.99) 0.86,1.09 −0.19 0.89 < 0.001 21 侧枝 30 1.16(1.13) 0.86,1.58 −0.58 0.42 < 0.001 9 30 1.83 1.43,2.35 −1.53 0.70 < 0.001 0 30 1.09(1.13) 0.82,1.45 −0.31 0.47 < 0.001
下载: 导出CSV
-
[1] 王飞,郭树江,纪永福,张莹花,韩福贵,等. 不同演替阶段白刺灌丛沙堆土壤因子与叶功能性状关系研究[J]. 干旱区地理,2022,45(1):176−184. Wang F,Guo SJ,Ji YF,Zhang YH,Han FG,et al. Relationship between soil factors and leaf functional traits of Nitraria tangutorum shrub at different succession stages[J]. Arid Land Geography,2022,45 (1):176−184.
[2] Cornelissen JHC,Lavorel S,Garnier E,Díaz S,Buchmann N,et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J]. Aust J Bot,2003,51 (4):335−380. doi: 10.1071/BT02124
[3] 李俊慧,彭国全,杨冬梅. 常绿和落叶阔叶物种当年生小枝茎长度和茎纤细率对展叶效率的影响[J]. 植物生态学报,2017,41(6):650−660. doi: 10.17521/cjpe.2016.0376 Li JH,Peng GQ,Yang DM. Effect of stem length to stem slender ratio of current-year twigs on the leaf display efficiency in evergreen and deciduous broadleaved trees[J]. Chinese Journal of Plant Ecology,2017,41 (6):650−660. doi: 10.17521/cjpe.2016.0376
[4] Westoby M,Wright IJ. Land-plant ecology on the basis of functional traits[J]. Trends Ecol Evol,2006,21 (5):261−268. doi: 10.1016/j.tree.2006.02.004
[5] 孙小妹,何明珠,周彬,李金霞,陈年来. 霸王根茎叶非结构性碳与C∶N∶P计量特征对干旱的响应[J]. 干旱区地理,2021,44(1):240−249. Sun XM,He MZ,Zhou B,Li JX,Chen NL. Non-structural carbohydrates and C∶N∶P stoichiometry of roots,stems,and leaves of Zygophyllum xanthoxylon in responses to xeric condition[J]. Arid Land Geography,2021,44 (1):240−249.
[6] 吴伟光,曹先磊. 毛竹林经营投入产出关系与经营效益分析[J]. 南京林业大学学报(自然科学版),2016,40(3):108−114. Wu WG,Cao XL. Study on the relationship between input,output and economic benefit of moso bamboo plantation[J]. Journal of Nanjing Forestry University (Natural Sciences Edition)
,2016,40 (3):108−114. [7] 姚钧能,吕建华,俞卫良,张君波,雷赵枫,等. 氮沉降和经营强度对毛竹林凋落叶生态化学计量特征的影响[J]. 应用生态学报,2018,29(2):467−473. Yao JN,Lü JH,Yu WL,Zhang JB,Lei ZF,et al. Effects of nitrogen deposition and management intensity on stoichiometry of leaf litter in Moso bamboo forest[J]. Chinese Journal of Applied Ecology,2018,29 (2):467−473.
[8] 冯鹏飞,李玉敏. 2021年中国竹资源报告[J]. 世界竹藤通讯,2023,21(2):100−103. [9] Busse MD,Cochran PH,Barrett JW. Changes in ponderosa pine site productivity following removal of understory vegetation[J]. Soil Sci Soc Am J,1996,60 (6):1614−1621. doi: 10.2136/sssaj1996.03615995006000060004x
[10] Hébert F,Thiffault N,Ruel JC,Munson AD. Ericaceous shrubs affect black spruce physiology independently from inherent site fertility[J]. Forest Ecol Manag,2010,260 (2):219−228. doi: 10.1016/j.foreco.2010.04.026
[11] Warton DI,Wright IJ,Falster DS,Westoby M. Bivariate line-fitting methods for allometry[J]. Biol Rev,2006,81 (2):259−291. doi: 10.1017/S1464793106007007
[12] Warton DI,Duursma RA,Falster DS,Taskinen S. SMART 3-an R package for estimation and inference about allometric lines[J]. Methods Ecol Evol,2012,3 (2):257−259. doi: 10.1111/j.2041-210X.2011.00153.x
[13] Warton DI,Weber NC. Common slope tests for bivariate errors-in-variables models[J]. Biometr J,2002,44 (2):161−174. doi: 10.1002/1521-4036(200203)44:2<161::AID-BIMJ161>3.0.CO;2-N
[14] 杨巧,朱润军,杨畅宇,李仕杰,程希平. 基于树形结构的木棉叶功能性状差异性研究[J]. 生态学报,2022,42(7):2834−2842. Yang Q,Zhu RJ,Yang CY,Li SJ,Cheng XP. Variation in leaf functional traits of Bombax ceiba Linnaeus communities based on tree structure[J]. Acta Ecologica Sinica,2022,42 (7):2834−2842.
[15] 曹永慧,周本智,葛晓改,倪霞,王小明. 毛竹比叶质量时空变化及对截雨干旱的响应[J]. 林业科学研究,2019,32(6):31−39. Cao YH,Zhou BZ,Ge XG,Ni X,Wang XM. Seasonal and canopy variation of leaf mass per area for Phyllostachys edulis leaves and its response to drought stress[J]. Forest Research,2019,32 (6):31−39.
[16] 邱东,吴甘霖,刘玲,涂文琴,陶冶. 城市香樟叶片干物质含量及比叶面积的时空变异[J]. 云南大学学报(自然科学版),2019,41(3):609−618. Qiu D,Wu GL,Liu L,Tu WQ,Tao Y. Spatial-temporal variation of leaf dry matter content and specific leaf area of Cinnamomum camphora in urban area[J]. Journal of Yunnan University (Natural Sciences Edition)
,2019,41 (3):609−618. [17] 戚德辉,温仲明,杨士梭,王红霞,郭茹. 基于功能性状的铁杆蒿对环境变化的响应与适应[J]. 应用生态学报,2015,26(7):1921−1927. Qi DH,Wen ZM,Yang SS,Wang HX,Guo R. Trait-based responses and adaptation of Artemisia sacrorum to environmental changes[J]. Chinese Journal of Applied Ecology,2015,26 (7):1921−1927.
[18] 刘希珍,封焕英,蔡春菊,范少辉,刘广路. 毛竹向阔叶林扩展过程中的叶功能性状研究[J]. 北京林业大学学报,2015,37(8):8−17. Liu XZ,Feng HY,Cai CJ,Fan SH,Liu GL. Response of leaf functional traits of Moso bamboo during the invading process into the broad-leaved forest[J]. Journal of Beijing Forestry University,2015,37 (8):8−17.
[19] 张运春,杜晓军,张桥英,高贤明,苏智先. 克隆乔木黄牛奶树枝条的功能特征[J]. 植物生态学报,2005,29(5):799−806. Zhang YC,Du XJ,Zhang QY,Gao XM,Su ZX. Functions of branches of the clonal tree Symplocos laurina[J]. Acta Phytoecologica Sinica,2005,29 (5):799−806.
[20] Day ME,Greenwood MS,Diaz-Sala C. Age- and size-related trends in woody plant shoot development:regulatory pathways and evidence for genetic control[J]. Tree Physiol,2002,22 (8):507−513. doi: 10.1093/treephys/22.8.507
[21] Thomas SC,Winner WE. Photosynthetic differences between saplings and adult trees:an integration of field results by meta-analysis[J]. Tree Physiol,2002,22 (2-3):117−127. doi: 10.1093/treephys/22.2-3.117
[22] 韩威,刘超,樊艳文,赵娜,叶思阳,等. 长白山阔叶木本植物叶片形态性状沿海拔梯度的响应特征[J]. 北京林业大学学报,2014,36(4):47−53. Han W,Liu C,Fan YW,Zhao N,Ye SY,et al. Responses of leaf morphological traits for broadleaved woody plants along the altitudinal gradient of Changbai Mountain,northeastern China[J]. Journal of Beijing Forestry University,2014,36 (4):47−53.
[23] 毛伟,李玉霖,张铜会,赵学勇,黄迎新,宋琳琳. 不同尺度生态学中植物叶性状研究概述[J]. 中国沙漠,2012,32(1):33−41. Mao W,Li YL,Zhang TH,Zhao XY,Huang YX,Song LL. Research advances of plant leaf traits at different ecology scales[J]. Journal of Desert Research,2012,32 (1):33−41.
[24] Long WX,Zang RG,Schamp BS,Ding Y. Within- and among-species variation in specific leaf area drive community assembly in a tropical cloud forest[J]. Oecologia,2011,167 (4):1103−1113. doi: 10.1007/s00442-011-2050-9
[25] Lohier T,Jabot F,Meziane D,Shipley B,Reich PB,Deffuant G. Explaining ontogenetic shifts in root-shoot scaling with transient dynamics[J]. Ann Bot,2014,114 (3):513−524. doi: 10.1093/aob/mcu128
[26] Moles AT,Westoby M. Do small leaves expand faster than large leaves,and do shorter expansion times reduce herbivore damage?[J]. Oikos,2000,90 (3):517−524. doi: 10.1034/j.1600-0706.2000.900310.x
[27] 朱旭斌,刘娅梅,孙书存. 南京地区落叶栎林主要木本植物的展叶动态研究[J]. 植物生态学报,2005,29(1):128−136. Zhu XB,Liu YM,Sun SC. Leaf expansion of the dominant woody species of three deciduous oak forests in Nanjing,east China[J]. Acta Phytoecologica Sinica,2005,29 (1):128−136.
[28] 宋利霞,陶建平,冉春燕,余小红,王永健,李媛. 卧龙亚高山暗针叶林不同林冠环境下华西箭竹的克隆生长[J]. 植物生态学报,2007,31(4):637−644. doi: 10.17521/cjpe.2007.0082 Song LX,Tao JP,Ran CY,Yu XH,Wang YJ,Li Y. Clonal growth of Fargesia nitida under different canopy conditions in a subalpine dark coniferous forest in Wolong nature reserve,China[J]. Journal of Plant Ecology ,2007,31 (4):637−644. doi: 10.17521/cjpe.2007.0082
[29] 莫丹,王振孟,左有璐,向双. 亚热带常绿阔叶林木本植物幼树阶段抽枝展叶的权衡关系[J]. 植物生态学报,2020,44(10):995−1006. doi: 10.17521/cjpe.2020.0143 Mo D,Wang ZM,Zuo YL,Xiang S. Trade-off between shooting and leaf developing of woody species saplings in subtropical evergreen broad-leaved forests[J]. Chinese Journal of Plant Ecology,2020,44 (10):995−1006. doi: 10.17521/cjpe.2020.0143
[30] Wang JW,Yu D,Wang Q. Growth,biomass allocation,and autofragmentation responses to root and shoot competition in Myriophyllum spicatum as a function of sediment nutrient supply[J]. Aquat Bot,2008,89 (4):357−364. doi: 10.1016/j.aquabot.2008.04.001
[31] Yu FH,Chen YF,Dong M. Clonal integration enhances survival and performance of Potentilla anserina,suffering from partial sand burial on Ordos plateau,China[J]. Evol Ecol,2001,15 (4-6):303−318. doi: 10.1023/A:1016032831038
[32] 朱强根,金爱武,王意锟,邱永华,李雪涛,张四海. 不同营林模式下毛竹枝叶的生物量分配:异速生长分析[J]. 植物生态学报,2013,37(9):811−819. Zhu QG,Jin AW,Wang YK,Qiu YH,Li XT,Zhang SH. Biomass allocation of branches and leaves in Phyllostachys heterocycla ‘Pubescens’ under different management modes:allometric scaling analysis[J]. Chinese Journal of Plant Ecology,2013,37 (9):811−819.
[33] 郭子武,杨清平,李迎春,陈双林. 密度对四季竹地上生物量分配格局及异速增长模式的制约性调节[J]. 生态学杂志,2013,32(3):515−521. Guo ZW,Yang QP,Li YC,Chen SL. Restrictive regulation of stand density on aboveground biomass allocation and allometric pattern of Oligostachyum lubricum[J]. Chinese Journal of Ecology,2013,32 (3):515−521.
[34] 施建敏,叶学华,陈伏生,杨清培,黎祖尧,等. 竹类植物对异质生境的适应——表型可塑性[J]. 生态学报,2014,34(20):5687−5695. Shi JM,Ye XH,Chen FS,Yang QP,Li ZY,et al. Adaptation of bamboo to heterogeneous habitat:phenotypic plasticity[J]. Acta Ecologica Sinica,2014,34 (20):5687−5695.
-
期刊类型引用(1)
1. 龙桂根,黄芝云,吴南生,冯胜,冯超,丁菲,金松松,何利人,王勇,陈玲. 南酸枣种质资源果实性状变异和综合评价. 热带亚热带植物学报. 2024(06): 781-790 .  百度学术
百度学术
其他类型引用(3)




计量
- 文章访问数: 170
- HTML全文浏览量: 42
- PDF下载量: 33
- 被引次数: 4
