
Identification and application of salt-tolerant germplasms at germination stage in foxtail millet(Setaria italica L.)
-
摘要:
本研究利用不同浓度NaCl溶液对10份谷子(Setaria italica L.)种质进行处理,通过分析其萌发期的相对发芽势、相对发芽率、相对芽长以及相对根长等4项指标,明确了适于谷子萌发期耐盐性鉴定的NaCl浓度为180 mmol/L。在该浓度下,利用主成分分析和聚类分析等方法,对180份种质资源进行了耐盐性综合评价和等级划分。结果显示,除相对发芽率和相对芽长之间相关性不显著以外,其余指标之间均呈极显著正相关;主成分分析结果表明,这4项指标可作为谷子耐盐性评价的重要指标;聚类分析结果将180份谷子种质分为极端耐盐、耐盐、盐敏感和极端盐敏感4类;进一步采用隶属函数进行综合评价,筛选到硷谷、衡谷12、齐头白、K-3606和晋谷20等5份极端耐盐种质材料。最后,在该浓度处理下,对黑枝谷×长农35号(极端盐敏感×耐盐)F7代重组近交系遗传群体进行了初步分析,发现40份株系耐盐性等级频率分布近似正态分布,表明该群体适宜耐盐QTL挖掘。研究结果说明,在180 mmol/L NaCl处理下,通过谷子萌发期相对发芽势、相对发芽率、相对芽长和相对根长等4个指标能较好地区分不同种质耐盐性的差异。
Abstract:In this study, 10 foxtail millet (Setaria italica L.) germplasm accessions were treated with varying concentrations of NaCl solution. By analyzing four indexes, including the relative germination potential, relative germination rate, relative shoot length, and relative root length during germination stage, we established that the suitable NaCl concentration for the identification of salt tolerance during germination in foxtail millet was 180 mmol/L. To evaluate the salt-tolerance of 180 core germplasms during the germination stage, correlation analysis and principal component analysis were employed to calculate the aforementioned four traits under 180 mmol/L NaCl treatment. Comprehensive evaluation and salt-tolerant grade of the 180 germplasms were identified by membership functional analysis and cluster analysis. Results revealed that except for the non-significant correlation between relative germination rate and relative shoot length, all other indexes showed a highly significant positive correlation. Furthermore, principal component analysis suggested that the four indexes could be considered as key indices for assessing salt tolerance. Cluster analysis revealed that the 180 germplasms could be divided into four groups: highly salt tolerant, salt tolerant, salt sensitive, and extremely salt sensitive. Jiangu, Henggu 12, Qitoubai, K-3606, and Jingu 20 were identified as extremely salt tolerant accessions through comprehensive evaluation using membership function. In addition, under 180 mmol/L NaCl stress, 40 lines from the F7 generation recombinant inbred line (RIL) of Heizhigu (extremely salt-sensitive)×Changnong 35 (salt-tolerant) were analyzed. Results showed significant variation among the 40 lines, with their frequency distribution of salt tolerance grades approximating normal distribution, indicating that this population was suitable for salt-tolerant QTL mapping. These findings demonstrated that 180 mmol/L NaCl could be used for the identification of salt-tolerant foxtail millet germplasms during the germination stage and that at this concentration, salt tolerance differences among various germplasms can be better distinguished by four indexes: relative germination potential, relative germination rate, relative shoot length, and relative root length.
-
Keywords:
- Setaria italica /
- Salt stress /
- Germination stage /
- Comprehensive evaluation
-
VQ(Valine-glutamine)是植物特有的一类蛋白,因其包含一个特有的保守基序FxxxVQxLTG而得名(F:苯丙氨酸,V:缬氨酸,Q:谷氨酰胺,L:亮氨酸,T:苏氨酸,G:甘氨酸,x 代表任意氨基酸)[1-3]。首个VQ蛋白SIB1(SIGMA FACTOR-BINDING PROTEIN1)在拟南芥(Arabidopsis thaliana (L.) Heynh.)中被发现[4],随后,从拟南芥中又陆续鉴定出33个VQ蛋白。目前,科研人员已在茶(Camellia sinensis (L.) O. Ktze. )[2]、大豆(Glycine max (L.) Merr.)[5]、小麦(Triticum aestivum L.)[6]及烟草(Nicotiana tabacum L.)[3]中分别鉴定出25、75、113和59个VQ基因。
VQ蛋白在植物应答生物与非生物胁迫中发挥着重要作用[7, 8],其作用机制主要有两种。一是通过与WRKY转录因子相互作用发挥功能,如AtVQ23和AtVQ16通过与WRKY33的WRKY结构域结合,增强后者与靶基因的结合能力,从而影响植物的抗病能力[9]。而AtVQ9与WRKY8互作抑制了WRKY8与W-box的结合活性,从而负调控植物的抗盐性[10]。在番茄(Solanum lycopersicum L.)叶片中,SlVQ7与SlWRKY37互作可提高后者的稳定性,并转录激活其下游靶基因,从而正调控茉莉酸及黑暗诱导的叶片衰老[11]。二是作为丝裂原活化蛋白激酶(MAPK)家族的磷酸化底物发挥作用[12-14]。如拟南芥MKS1(AtVQ21)作为AtMPK4的底物,与WRKY25和WRKY33相互作用,有助于激活MPK4调节的病原体防御反应[3]。
多年生黑麦草(Lolium perenne L.)为禾本科早熟禾亚科黑麦草属植物,是世界上广泛种植的冷季型草坪草及牧草[15],具有分蘖多、成坪速度快、粗蛋白含量高、耐牧性好等优点。多年生黑麦草在生长发育过程中常面临高温、盐碱、荫蔽等非生物胁迫,严重影响草坪质量和牧草品质。因此,鉴定多年生黑麦草VQ基因,探究其在应答非生物胁迫中的作用十分必要。然而,目前尚未见相关报道。本研究系统鉴定了多年生黑麦草VQ蛋白家族成员,并对其保守结构域、进化关系、共线性、亚细胞定位及其在高温、盐碱、黑暗等非生物胁迫下的表达模式进行了分析,研究结果旨在为进一步探讨VQ蛋白在多年生黑麦草生长发育及应答非生物逆境胁迫中的作用奠定基础。
1. 材料与方法
1.1 实验材料
本研究以多年生黑麦草品种‘百灵鸟’为实验材料。黑麦草种子于营养土(基质∶蛭石=1∶1)中萌发,萌发后转移至光照培养箱,光周期为16 h / 8 h(光 / 黑暗),温度为25 ℃ / 22 ℃(光 / 黑暗)。待植株生长1个月后进行黑暗、高温和NaCl处理。黑暗处理的条件为:25 ℃ / 22 ℃,24 h黑暗,分别于处理0、2、8 d后取样。高温处理的条件为:42 ℃,光周期16 h / 8 h,分别于处理0、1、24 h后取样。NaCl处理的条件为:25 ℃ / 22 ℃,光周期16 h / 8 h,使用200 mL 200 mmol/L的NaCl溶液进行浇灌,分别于盐处理0、3、38 h后取样。
1.2 多年生黑麦草VQ基因家族成员的鉴定
以Pfam数据库中VQ结构域(PF05678)的隐马尔可夫模型(HMM)图谱作为查询,通过HMMER搜索多年生黑麦草基因组,鉴定了可能的VQ蛋白序列。使用Pfam数据库(http://pfam.xfam.org/)进一步确认了这些序列。在拟南芥基因组数据库(TAIR)中下载拟南芥全基因组序列及其注释信息。
1.3 多年生黑麦草VQ家族成员的理化性质及亚细胞定位
使用在线网站(https://web.expasy.org/compute_pi/)分析多年生黑麦草VQ蛋白的理化性质。使用BIOINF(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-)在线工具对多年生黑麦草VQ家族成员进行亚细胞定位预测。
1.4 多年生黑麦草VQ家族基因系统发育树构建
使用MEGAX(http://www.megasoftware.net)软件的ML法构建系统发育树,进行系统发育分析,并用iTOL(https://itol.embl.de/)网站进行美化。
1.5 多年生黑麦草VQ保守基序及基因结构分析
利用MEME(https://meme-suite.org/meme/tools/meme)网站对VQ蛋白的氨基酸序列进行保守基序分析。结合植物基因组序列和注释文件,用TBtools提取VQ基因的基因组序列及CDS序列,用TBtools软件工具中的Gene Structure View进行可视化。
1.6 VQ基因染色体分布及种内共线性分析
结合植物基因组序列和注释文件,使用TBtools软件工具中的Amazing Gene Location From GFF3/GTF File进行可视化,使用TBtools软件工具中的one step MCScanX进行种内基因组比对,并用Multiple Synreny Plot进行可视化。
1.7 胁迫条件下多年生黑麦草VQ基因的表达模式分析
利用Trizol试剂(Takara,大连)提取多年生黑麦草在不同非生物胁迫处理后的总RNA。使用反转录试剂盒Hifair® Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus)(Yeasen,上海)合成cDNA。利用Hieff® qPCR SYBR Green Master Mix(High Rox Plus)试剂盒(Yeasen,上海)在Step One Plus Real-Time PCR Systems系统上进行qRT-PCR反应。反应体系总体积为20 μL,包括2 μL cDNA,上、下游引物各0.8 μL,10 μL SYBR,用ddH2O补足体积。反应程序为:95 ℃预变性5 min;95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸20 s,40个循环,系统自带熔解曲线。引物序列见附表1
1 ,由擎科公司(擎科,武汉)合成,多年生黑麦草eEF1A(s)为内参基因。采用2−△△Ct方法计算候选基因的相对表达量,实验设3次重复。利用TBtools绘制热图,进行表达模式分析。2. 结果与分析
2.1 LpVQ家族成员鉴定
为了系统分析多年生黑麦草的VQ基因,以VQ结构域(PF05678)为参考,从全基因组水平挖掘其VQ家族基因。结果显示,多年生黑麦草中共存在52个VQ基因。对这些基因的编码产物LpVQ家族成员进行理化性质分析,发现LpVQs的氨基酸长度介于118~427 aa,蛋白分子量范围为11.67~43.84 kD,等电点在5.01~11.6。通过BIOINF在线网站对LpVQ家族成员进行亚细胞定位预测,发现48个蛋白定位在细胞核中,LpVQ22为叶绿体与细胞核共定位,LpVQ33为细胞壁与细胞核共定位,LpVQ37为细胞膜与细胞核共定位,而LpVQ45预测定位在细胞膜中(附表2
1 )。2.2 LpVQ家族系统进化树分析
为了分析LpVQs的进化关系,利用多年生黑麦草的52个LpVQ蛋白和拟南芥的34个AtVQ蛋白共同构建系统进化树。结果显示,拟南芥与多年生黑麦草的VQ家族成员可划分为7个亚家族(Ⅰ~Ⅶ),分别包含5、8、6、11、12、6和4个LpVQ蛋白。亚家族Ⅰ中只包含LpVQs,亚家族Ⅳ、Ⅴ和Ⅵ中含有较多的LpVQ家族成员,而AtVQ家族成员则更多地归类于亚家族Ⅱ、Ⅲ和Ⅶ(图1)。进一步分析发现,多年生黑麦草与拟南芥的VQ家族间存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。
![]() 图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne
图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne2.3 LpVQ家族成员保守基序和基因结构分析
为了研究蛋白的结构多样性,利用MEME在线软件对52个LpVQ蛋白的保守基序进行分析,结果发现17%的LpVQs含有1个Motif,48%的LpVQs含有两个Motif,19%的LpVQs含有3个Motif,6%的LpVQs含有4个Motif,10%的LpVQs含有5个Motif。所有家族成员均具有Motif1,表明它是LpVQ家族的核心保守结构域(图2)。
此外,本研究用TBtools工具对LpVQs进行了基因结构分析。结果发现,46个LpVQ基因没有内含子,5个LpVQ基因含有1个内含子(LpVQ24、LpVQ29、LpVQ31、LpVQ32、LpVQ52),仅有1个基因(LpVQ161)含有两个内含子(附图1
1 )。2.4 LpVQ家族成员染色体定位以及共线性分析
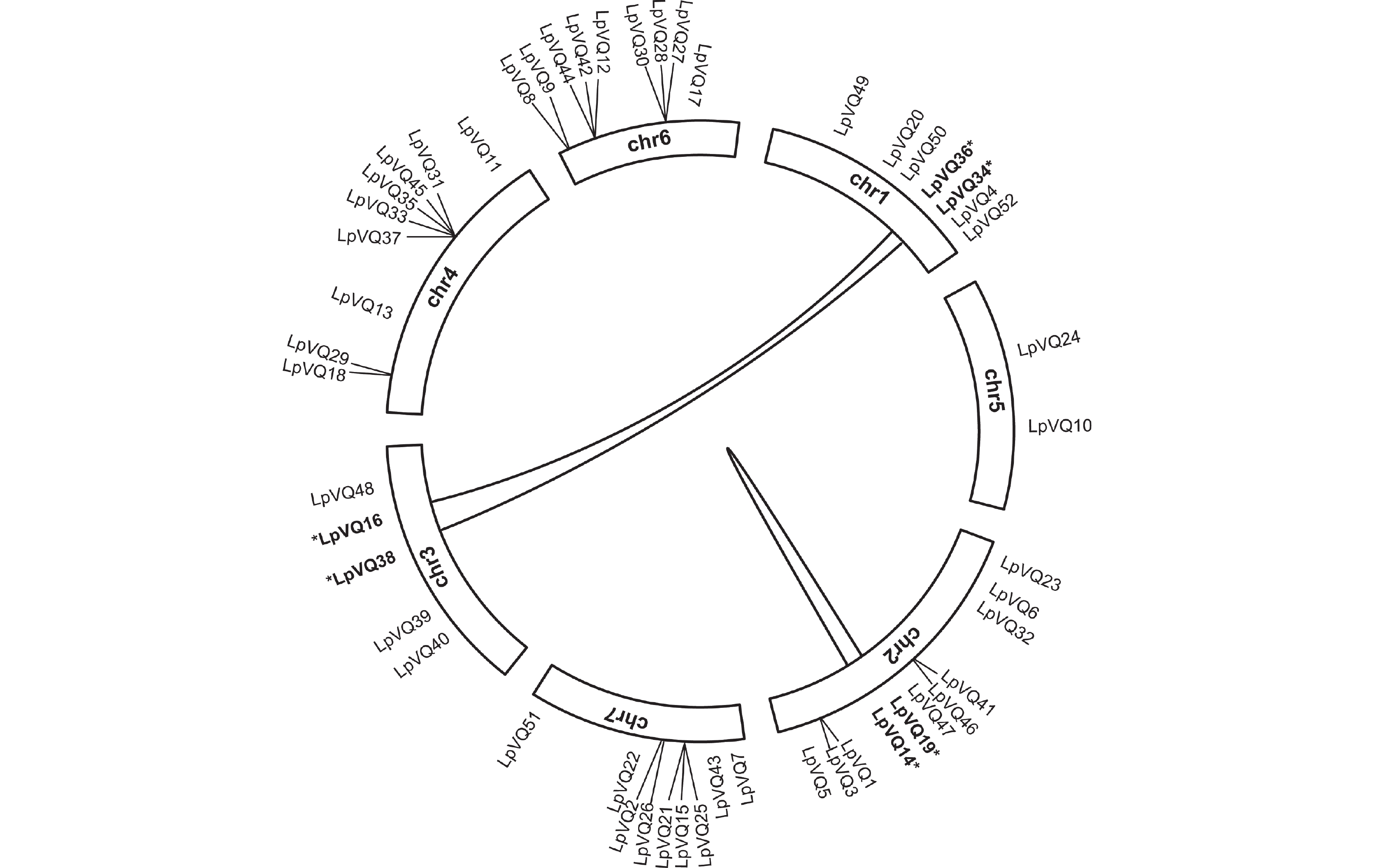
染色体定位分析结果表明,LpVQ基因分布在多年生黑麦草的7条染色体上,其中分布在chr1上的有7个,占13%,分布在chr2上的最多,为11个,占21%,分布在chr3上的为5个,占10%,分布在chr4、chr6和chr7上的基因均各有9个,各占17%,而分布在chr5上的最少,仅有2个,占4%(图3)。
为了了解LpVQ基因之间的进化关系,本研究对VQ家族成员进行了种内共线性分析。结果表明,3对VQ基因之间存在共同进化关系,分别是LpVQ16与LpVQ36、LpVQ34与LpVQ38以及LpVQ14与LpVQ19(图4)。
2.5 LpVQ基因响应非生物胁迫的表达模式分析
从7个亚家族中分别随机选取3个LpVQ基因,检测其在黑暗、高温以及盐胁迫下多年生黑麦草中的表达量变化。结果显示,黑暗处理后,只有4个LpVQ基因的表达持续上调。大部分LpVQ基因的表达量在黑暗处理2 d后下降,其中,LpVQ2、LpVQ8、LpVQ14、LpVQ22及LpVQ38的表达量在黑暗处理8 d后又显著上升(图5:A)。
![]() 图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.
图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.42 ℃高温处理后,LpVQ的表达模式被分为3类。LpVQ18、LpVQ8、LpVQ28、LpVQ2、LpVQ7、LpVQ4及LpVQ12受1 h高温处理的显著诱导,而高温处理24 h后,其表达又显著下调。LpVQ9、LpVQ22、LpVQ15、LpVQ3、LpVQ42、LpVQ6、LpVQ10、LpVQ1、LpVQ38的表达量在1 h和24 h高温处理后均显著下降。LpVQ5、LpVQ27、LpVQ34、LpVQ14和LpVQ44的表达量只在24 h高温处理后下降(图5:B)。
NaCl处理3 h后,13个LpVQ基因显著上调表达,其中LpVQ4的表达上升了13倍。而NaCl处理38 h后,大部分基因的表达明显下调。仅有3个基因的表达受盐诱导持续上调,分别是 LpVQ8、LpVQ10和LpVQ38(图5:C)。
3. 讨论
近年来,VQ基因已经在多种植物中被发掘鉴定,包括拟南芥、水稻(Oryza sativa L.)、大豆和葡萄(Vitis vinifera L.)等[16-19],而关于多年生黑麦草VQ家族基因的鉴定和分析还鲜见报道。本研究通过全基因组鉴定,得到52个多年生黑麦草LpVQ基因,并系统分析了其理化特性、蛋白结构、基因结构、共线性、进化关系以及响应逆境胁迫的表达模式。
基因结构分析发现,88.46%的LpVQ基因无内含子,与之类似,在拟南芥[16]、水稻[17]、玉米(Zea mays L.)[20]和马铃薯(Solanum tuberosum L.)[21]中,无内含子的VQ基因分别占88.2%、92.5%、88.5%和91.89%。而苔藓(Bryophyta)[22]中无内含子的VQ基因比例则远低于以上物种,仅为28.0%,说明VQ基因可能在长期进化过程中丢失了内含子。
系统进化分析结果显示,多年生黑麦草和拟南芥的VQ家族成员可被分为7个亚家族,且LpVQs与AtVQs亲缘关系较远,只存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。根据拟南芥VQ基因的功能,可以推测多年生黑麦草中其直系同源VQ基因的功能。有研究指出,AtVQ9作为AtWRKY8的抑制子,能提高拟南芥的耐盐性[9],由此推测,其直系同源基因LpVQ11可能也在多年生黑麦草抵御盐胁迫中发挥重要作用。
VQ基因在植物响应非生物胁迫中发挥着重要作用。如,过表达PtVQ1基因显著增强了转基因拟南芥的耐盐性[23];而过表达MdVQ37则提高了转基因苹果(Malus pumila Mill.)对高温胁迫的敏感性[24]。本研究对黑暗、高温及盐胁迫处理下多年生黑麦草的VQ表达量进行了分析。结果显示,LpVQ基因广泛响应黑暗、高温及盐胁迫,且大部分LpVQ基因均在胁迫初期即做出快速响应,这与西葫芦(Cucurbita pepo L.)[8]、黄瓜(Cucumis sativus L.)[25]等物种中的研究结果相类似。然而,LpVQ基因的表达模式在不同胁迫处理下也存在差异。如,LpVQ1在黑暗胁迫处理后持续下调表达,而在高温和盐胁迫下持续上调表达,推测该基因可能在不同胁迫中发挥着不同的作用。尽管LpVQ可能参与多年生黑麦草对黑暗、高温及盐等胁迫响应的调控,但其具体功能还需要进一步验证。
综上所述,本研究对多年生黑麦草VQ基因家族成员进行了系统鉴定与分析,为后续验证VQ家族成员在多年生黑麦草应答生物或非生物胁迫中的功能提供了基础。
1 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。2 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。3 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
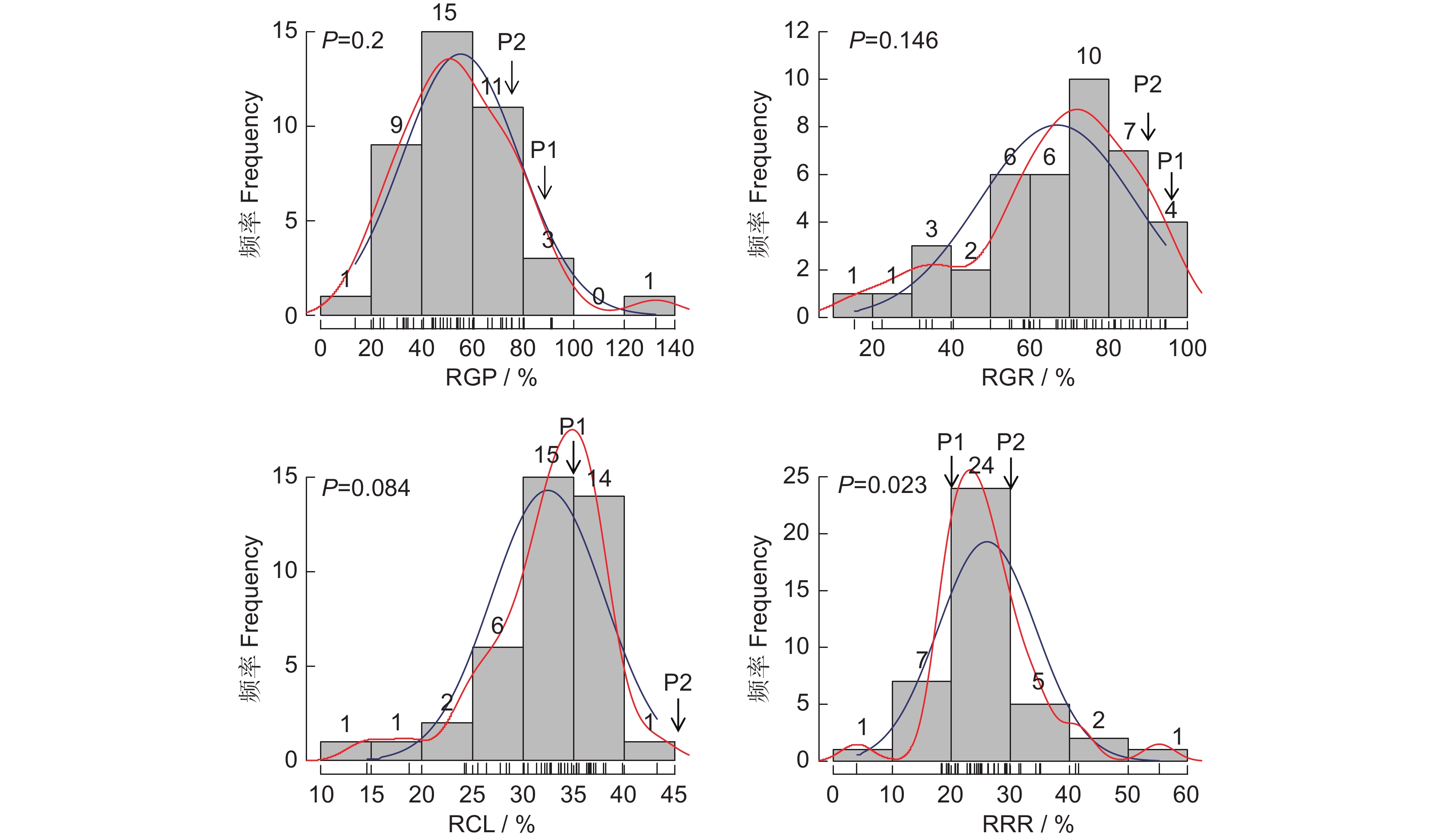
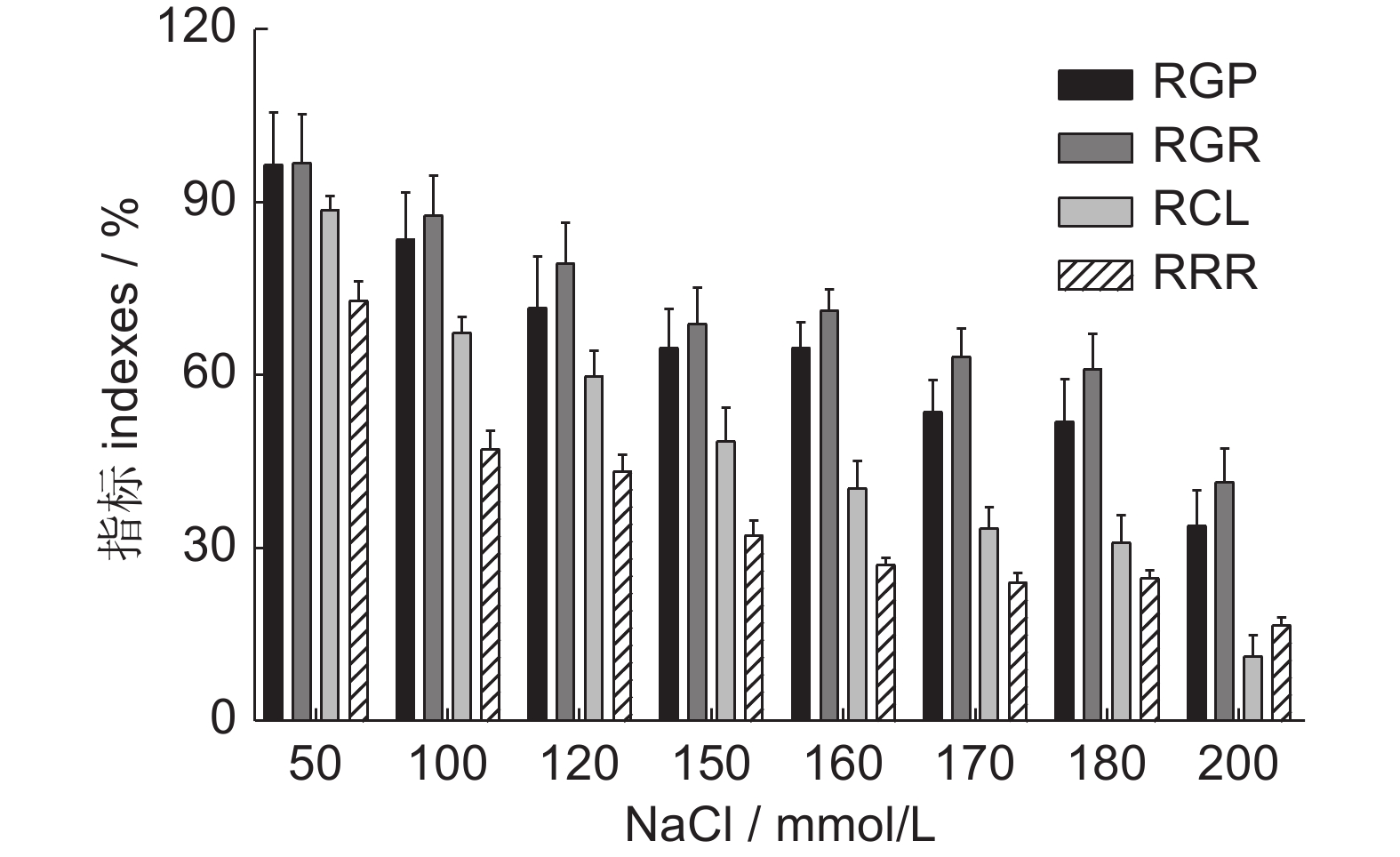
图 1 不同浓度NaCl处理下的4个指标分析
RGP:相对发芽势;RGR:相对发芽率;RCL:相对芽长;RRR:相对根长。下同。
Figure 1. Analysis of four indexes under different NaCl treatments
RGP: Relative germination potential; RGR: Relative germination rate; RCL: Relative coleoptile length; RRR: Relative radical root length. Same below.
![]()
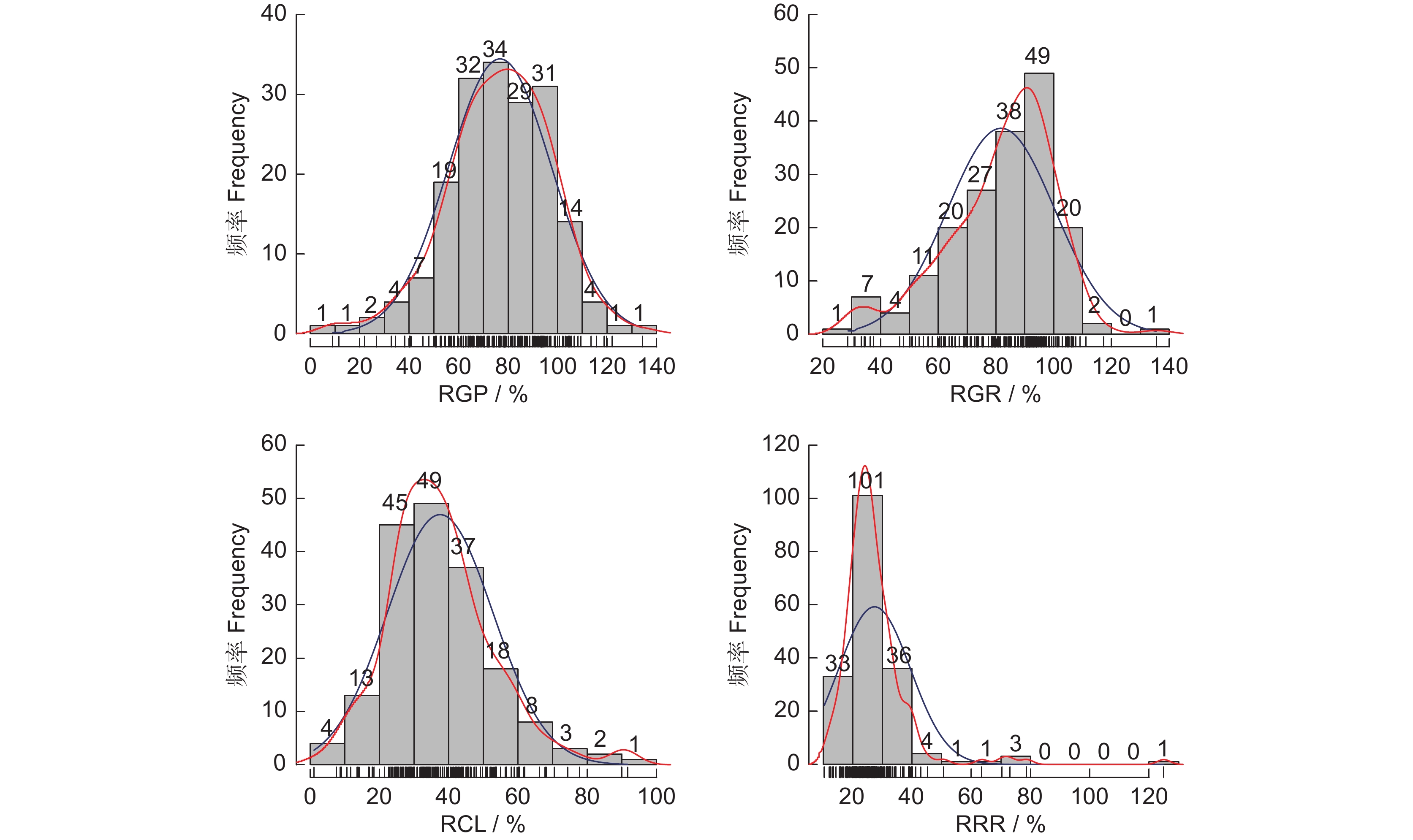
图 2 谷子萌发期耐盐性指标频数分布直方图
横坐标表示相对值大小,纵坐标表示各组所占频数,图中红线表示密度线,蓝线表示正态分布曲线。
Figure 2. Frequency distribution of salt tolerance traits during thegermination stage in Setaria italica
Abscissa represents relative value; Ordinate represents frequency of each group; Red line represents density; Blue line represents normal distribution curve.
![]()
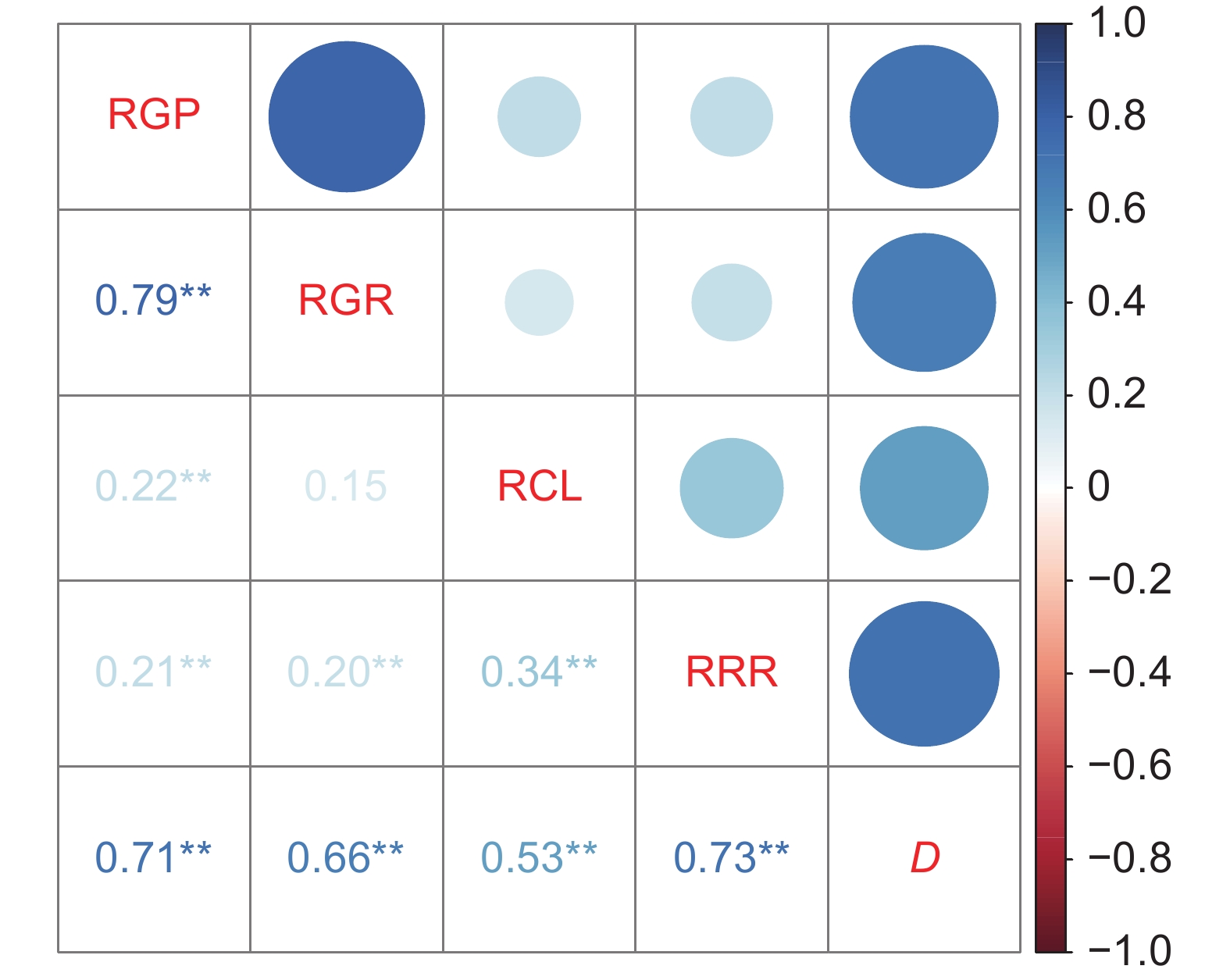
图 3 NaCl胁迫条件下谷子萌发期各指标相对值的相关性分析
D:综合评价值;**:表示在 0.01 水平上显著相关。
Figure 3. Correlation analysis of four indexes during the germination stage under NaCl stress in Setaria italica
D: Comprehensive evaluation value; **: Values with different superscript indicate significant different at 1% level.
![]()
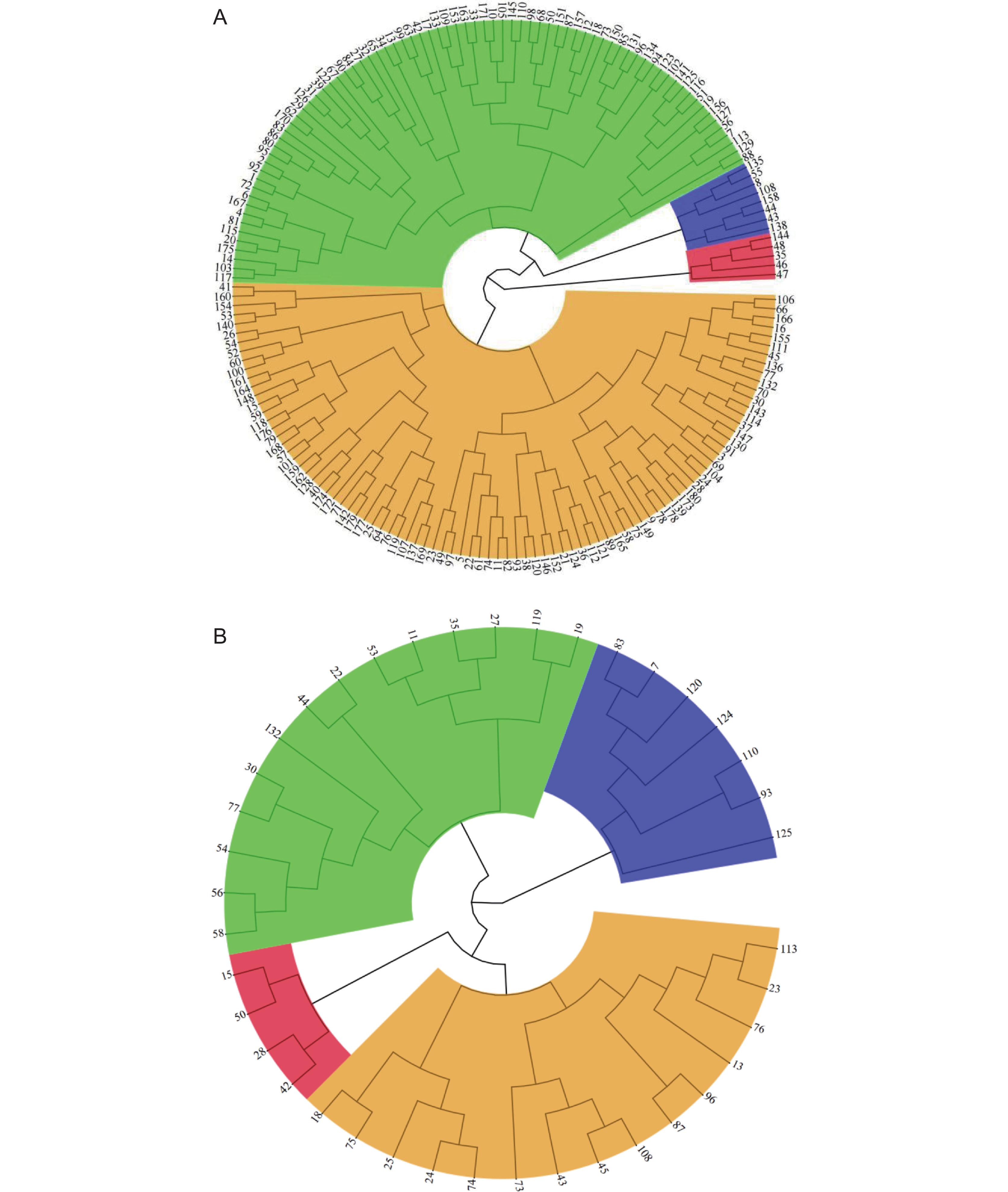
图 4 180份谷子种质及40份遗传群体后代耐盐性的聚类分析
A:180份谷子种质材料;数字代表180份谷子种质的编号(附表3)。B:遗传群体40份株系;数字代表RIL群体40份种质的编号;红色代表极端耐盐种质;黄色代表耐盐种质;绿色代表盐敏感种质;蓝色代表极端盐敏感种质。
Figure 4. Cluster of 180 germplasms and 40 lines from one genetic population for salt tolerance
A: 180 germplasms; Numbers represent codes of 180 germplasms(Attached Table 3). B: 40 lines from one genetic population; Numbers represent the code of 40 germplasms from a RIL population. Red represents extremely salt-tolerant accessions; Yellow represents salt-tolerant accessions; green represents salt-sensitive accessions; Blue represents extremely salt-sensitive accessions.
![]()
表 1 10份供试种质材料及来源
Table 1 Name and source of 10 germplasms
编号Code 名称Name 来源Origin 1 长农35号 山西 2 六月鲜 河北 3 龙辐 黑龙江 4 毛毛谷 河北 5 小乌谷 河北 6 金棒子 甘肃 7 茶清谷 河北 8 小红谷 甘肃 9 菠菜腿 内蒙古 10 郑448 河南  下载: 导出CSV
下载: 导出CSV
表 2 10份谷子种质在不同盐浓度下的指标分析
Table 2 Analysis of four indexes in 10 germplasms of Setaria italica under different NaCl concentrations
指标
IndexesNaCl浓度
NaCl
concentration /
mmol/L性状值
Average±
SE / %变异幅度
Range of
variation / %变异系数
CV / %RGP 50 96.58±27.64a 58.70~166.67 28.62 100 83.63±23.98ab 57.50~138.10 28.67 120 71.79±26.69b 45.00~139.68 37.18 150 64.88±20.19b 37.59~96.83 31.12 160 64.88±13.16b 54.00~91.00 20.29 170 53.63±16.72bc 33.00~87.00 31.17 180 52.07±25.58b 10.00~94.00 49.12 200 34.00±18.26c 10.00~67.44 53.70 RGR 50 96.91±25.90a 66.41~155.56 26.72 100 87.70±21.10ab 54.10~127.78 24.06 120 79.42±21.00 ab 56.10~134.72 26.51 150 68.97±19.54b 39.34~101.39 28.34 160 71.22±11.21b 61.00~96.00 15.74 170 63.37±14.32b 40.00~90.00 22.60 180 61.10±24.46b 12.00~89.00 40.03 200 41.52±17.36c 6.56~63.16 41.80 RCL 50 88.61±8.69a 73.87~101.19 9.81 100 67.40±9.88b 50.70~81.92 14.66 120 59.83±13.85b 37.38~82.34 23.15 150 48.53±18.03c 16.59~68.66 37.15 160 40.42±14.27cd 14.00~62.00 35.30 170 33.49±11.18d 12.00~54.00 33.38 180 31.07±16.85d 2.00~55.00 54.23 200 11.37±10.67e 0.00~32.15 93.87 RRR 50 72.93±11.22c 59.21~94.54 15.38 100 47.25±9.43b 31.94~57.26 19.97 120 43.36±8.07b 28.44~51.31 18.62 150 32.30±9.63c 18.40~49.43 29.80 160 27.10±3.77c 23.00~33.00 13.89 170 24.18±4.41c 18.00~34.00 18.23 180 24.82±5.65c 13.00~34.00 22.74 200 16.79±4.06d 9.72~24.99 24.17 注:指标名缩写同图1。同一指标同列不同字母表示不同处理间在 P<0.05水平上差异显著。 Notes: Abbreviations are the same as those in Fig. 1. Different letters indicate significant differences between treatments (P<0.05).
下载: 导出CSV
表 3 NaCl 胁迫下谷子萌发期综合指标的主成分及贡献率
Table 3 Principal component and contribution rates of comprehensive indexes of Setaria italica during the germination stage under NaCl stress
主成分
Principal component特征值
Eigen value贡献率
Contribution rate / %累计贡献率
Cumulative contribution rate / %1 2.046 51.140 51.140 2 1.107 27.687 78.827
下载: 导出CSV
表 4 NaCl胁迫条件下谷子萌发期各指标的因子载荷矩阵
Table 4 Factor loading matrix of four indexes of Setaria italica during the germination stage under NaCl stress
指标
Indexes成分1
Component 1成分2
Component 2RGP 0.439 −0.293 RGR 0.425 −0.353 RCL 0.260 0.540 RRR 0.218 0.633 注:指标名缩写同图1。 Note: Abbreviations are the same as those given in Fig. 1.
下载: 导出CSV
表 5 180份谷子种质萌发期耐盐分类情况
Table 5 Classification of 180 Setaria italica germplasms for salt tolerance during the germination stage
耐盐性
Salt tolerance综合评价值
D种质名称
Name极端耐盐 0.335~0.436 硷谷、衡谷12、齐头白、K-3606、晋谷20 耐盐 0.200~0.311 长治204、济矮秆6、小黄谷子、晋谷33、大青苗、小三变、小白谷、长农35号等 盐敏感 0.121~0.199 红苗金耙齿、鸭子嘴、显256B、金香玉、高粱红、黄粘谷、六棱谷、猫屎谷等 极端盐敏感 0.058~0.113 小青谷、黑谷、Red manna、秃头、锤罗、蒙金谷1号、山东-4、黑枝谷
下载: 导出CSV
-
[1] 王遵亲. 中国盐渍土[M]. 北京: 科学出版社, 1993: 400-515. [2] 中国科学院国家计划委员会, 自然资源考察委员会. 《中国1∶100万土地资源图》土地资源数据集[M]. 北京: 中国人民大学出版社, 1991: 89-175. [3] 全国土壤普查办公室. 中国土壤[M]. 北京: 中国农业出版社, 1998: 1253. [4] 马子林,马玉兰. 盐碱地的形成原因及改良措施[J]. 青海农牧业,2007(2):18. [5] 黄琼. 浅谈盐碱地的改良措施与对策[J]. 现代园艺,2016(22):230−231. [6] 殷炳政,张怀东,周彬. 农业综合开发治理盐碱地措施和方法初探[J]. 山东省农业管理干部学院学报,2013,30(4):38−39. [7] 袁杰,王学强,张燕红,赵志强,贾春平,等. 水稻种质资源苗期耐盐性鉴定[J]. 分子植物育种,2020,18(20):6808−6814. Yuan J,Wang XQ,Zhang YH,Zhao ZQ,Jia CP,et al. Identification and screening of salt tolerance in rice germplasm resources at seedling stage[J]. Molecular Plant Breeding,2020,18(20):6808−6814.
[8] 李小康,吴崇宁,王维,李文淑,Masahiro K,等. 人工合成六倍体小麦耐盐种质资源的筛选及评价[J]. 麦类作物学报,2021,41(12):1487−1495. Li XK,Wu CN,Wang W,Li WS,Masahiro K,et al. Screening and evaluation of salt-tolerant germplasm of synthetic hexaploidy wheat[J]. Journal of Triticeae Crops,2021,41(12):1487−1495.
[9] 段雅娟,曹士亮,于滔,李文跃,杨耿斌,等. 玉米自交系萌发期耐盐性鉴定[J]. 作物杂志,2022(1):213−219. Duan YJ,Cao SL,Yu T,Li WY,Yang GB,et al. Identification of salt tolerance during germination of maize inbred lines[J]. Crops,2022(1):213−219.
[10] 胡亮亮,王素华,王丽侠,程须珍,陈红霖. 绿豆种质资源苗期耐盐性鉴定及耐盐种质筛选[J]. 作物学报,2022,48(2):367−379. doi: 10.3724/SP.J.1006.2022.04283 Hu LL,Wang SH,Wang LX,Cheng XZ,Chen HL. Identification of salt tolerance and screening of salt tolerant germplasm of mungbean (Vigna radiate L. ) at seedling stage[J]. Acta Agronomica Sinica,2022,48(2):367−379. doi: 10.3724/SP.J.1006.2022.04283
[11] 张海金. 谷子在旱作农业中的地位和作用[J]. 安徽农学通报,2007,13(10):169−170. [12] 张一中,张一弓,柳青山. 谷子在山西省旱作农业中的地位和作用[J]. 中国种业,2011(8):21−22. [13] 刁现民. 中国谷子生产与发展方向[M]//柴岩, 万福世, 编. 中国小杂粮产业发展报告. 北京: 中国农业出版社, 2007: 32-43. [14] 韩飞,诸葛玉平,娄燕宏,王会,张乃丹,等. 63份谷子种质的耐盐综合评价及耐盐品种筛选[J]. 植物遗传资源学报,2018,19(4):685−693. Han F,Zhuge YP,Lou YH,Wang H,Zhang ND,et al. Evaluation of salt tolerance and screening for salt tolerant accessions of 63 foxtail millet germplasm[J]. Journal of Plant Genetic Resources,2018,19(4):685−693.
[15] 秦岭,张艳亭,陈二影,杨延兵,黎飞飞,管延安. 谷子萌发期耐盐种质筛选及其保护酶系统对盐胁迫的响应[J]. 中国农业科学,2019,52(22):4027−4038. Qin L,Zhang YT,Chen EY,Yang YB,Li FF,Guan YA. Screening for germplasms tolerant to salt at germination stage and response of protective enzymes to salt stress in foxtail millet[J]. Scientia Agricultura Sinica,2019,52(22):4027−4038.
[16] 张笛. 谷子耐盐种质资源筛选及生长发育与农艺性状的鉴定[D]. 黑龙江: 黑龙江八一农垦大学, 2020: 5-60. [17] 田伯红,王素英,李雅静,王建广,张立新,等. 谷子地方品种发芽期和苗期对NaCl胁迫的反应和耐盐品种筛选[J]. 作物学报,2008,34(12):2218−2222. Tian BH,Wang SY,Li YJ,Wang JG,Zhang LX,et al. Response to sodium chloride stress at germination and seedling and identification of salinity tolerant genotypes in foxtail millet landraces originated from China[J]. Acta Agronomica Sinica,2008,34(12):2218−2222.
[18] 张笛,苗兴芬,王雨婷. 100份谷子品种资源萌发期耐盐性评价及耐盐品种筛选[J]. 作物杂志,2019(6):43−49. Zhang D,Miao XF,Wang YT. Evaluation and screening of salt tolerance in 100 foxtail millet at germination stage[J]. Crops,2019(6):43−49.
[19] 谢志坚. 农业科学中的模糊数学方法[M]. 武汉: 华中理工大学出版社, 1983: 99-193. [20] 戴海芳,武辉,阿曼古丽·买买提阿力,王立红,麦麦提·阿皮孜,等. 不同基因型棉花苗期耐盐性分析及其鉴定指标筛选[J]. 中国农业科学,2014,47(7):1290−1300. Dai HF,Wu H,Amanguli · Maimaitiali,Wang LH,Maimaiti · Apizi,et al. Analysis of salt-tolerance and determination of salt-tolerant evaluation indicators in cotton seedlings of different genotypes[J]. Scientia Agricultura Sinica,2014,47(7):1290−1300.
[21] Khan AA,Rao SA,McNeilly T. Assessment of salinity tolerance based upon seedling root growth response functions in maize (Zea mays L.)[J]. Euphytica,2003,131(1):81−89. doi: 10.1023/A:1023054706489
[22] Verma OPS,Yadava RBR. Salt tolerance of some oats (Avena sativa L.) varieties at germination and seedling stage[J]. J Agron Crop Sci,1986,156(2):123−127. doi: 10.1111/j.1439-037X.1986.tb00016.x
[23] 智慧,刁现民,吕芃,李伟,Akolavo Z. 人工盐胁迫法鉴定谷子及狗尾草物种耐盐基因型[J]. 河北农业科学,2004,8(4):15−18. Zhi H,Diao XM,Lü P,Li W,Akolavo Z. Methodology analysis on screening of salt tolerant genotypes from foxtail millet and other Setaria species[J]. Journal of Hebei Agricultural Sciences,2004,8(4):15−18.
[24] 崔兴国,时丽冉. 衡水地区14份夏谷品种种子萌发期耐盐性研究[J]. 作物杂志,2011(4):117−119. Cui XG,Shi LR. Evaluation of salt tolerance of 14 foxtail millet summer varieties in Hengshui[J]. Crops,2011(4):117−119.
[25] 田伯红,王建广,李雅静,张立新,孔德平. 谷子发芽期和幼苗前期耐盐性鉴定指标的研究[J]. 河北农业科学,2008,12(7):4−6. Tian BH,Wang JG,Li YJ,Zhang LX,Kong DP. Studies on screening indexes for salt tolerance of millet during germination period and early seedling stage[J]. Journal of Hebei Agricultural Sciences,2008,12(7):4−6.
[26] Ardie SW,Khumaida N,Nur A,Fauziah N. Early identification of salt tolerant foxtail millet (Setaria italica L. Beauv)[J]. Proc Food Sci,2015,3:303−312. doi: 10.1016/j.profoo.2015.01.033
[27] Pan JW,Li Z,Dai SJ,Ding HF,Wang QG,et al. Integrative analyses of transcriptomics and metabolomics upon seed germination of foxtail millet in response to salinity[J]. Scientific Reports,2020,10(1):13660. doi: 10.1038/s41598-020-70520-1
[28] 陈二影,秦岭,杨延兵,黎飞飞,王润丰,等. 生产条件下谷子品种盐碱耐性的差异及综合评价[J]. 中国农业科学,2019,52(22):4050−4065. Chen EY,Qin L,Yang YB,Li FF,Wang RF,et al. Variation and comprehensive evaluation of salt and alkali tolerance of different foxtail millet cultivars under production conditions[J]. Scientia Agricultura Sinica,2019,52(22):4050−4065.
[29] 张艳亭. 谷子种质资源耐盐性鉴定和耐盐机理的研究[D]. 曲阜: 曲阜师范大学, 2018: 7-9. [30] 郭瑞锋,张永福,任月梅,杨忠. 混合盐碱胁迫对谷子萌发、幼芽生长的影响及耐盐碱品种筛选[J]. 作物杂志,2017(4):63−66. Guo RF,Zhang YF,Ren YM,Yang Z. Effects of saline-alkali stress on millet germination and shoots growth and saline-alkali tolerance variety screening[J]. Crops,2017(4):63−66.
[31] 陈二影,王润丰,秦岭,杨延兵,黎飞飞,等. 谷子芽期耐盐碱综合鉴定及评价[J]. 作物学报,2020,46(10):1591−1604. Chen EY,Wang RF,Qin L,Yang YB,Li FF,et al. Comprehensive identification and evaluation of foxtail millet for saline-alkaline tolerance during germination[J]. Acta Agronomica Sinica,2020,46(10):1591−1604.
[32] 虞晓芬,傅玳. 多指标综合评价方法综述[J]. 统计与决策,2004(11):119−121. [33] 张巧凤,陈宗金,吴纪中,蒋彦婕,杨继书,等. 小麦种质芽期和苗期的耐盐性鉴定评价[J]. 植物遗传资源学报,2013,14(4):620−626. Zhang QF,Chen ZJ,Wu JZ,Jiang YJ,Yang JS,et al. Screening for salinity tolerance at germination and seedling stages in wheat germplasm[J]. Journal of Plant Genetic Resources,2013,14(4):620−626.
[34] 周和平,张立新,禹锋,李平. 我国盐碱地改良技术综述及展望[J]. 现代农业科技,2007(11):159−161. [35] Ren ZH,Gao JP,Li LG,Cai XL,Huang W,et al. A rice quantitative trait locus for salt tolerance encodes a sodium transporter[J]. Nat Genet,2005,37(10):1141−1146. doi: 10.1038/ng1643
[36] Niones JM. Five mapping of the salinity tolerance gene on chromosome 1 of rice (Oryza sativa L.) using near-isogenic lines[M]. Laguna: University of the Philippines, 2004: 1-100.
-
其他相关附件
-
PDF格式
吕宗环附表1-3 点击下载(690KB)
-




计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
