
Regulation of hypocotyl elongation in Arabidopsis thaliana (L.) Heynh. by U2BL
-
摘要:
下胚轴的伸长在植物早期存活和后期生长发育过程中均具有重要作用。本研究对拟南芥(Arabidopsis thaliana (L.) Heynh.)突变体进行了筛选,得到了具有短下胚轴表型的突变体u2bl,并对其下胚轴变短机制进行了初步研究。结果显示,突变体u2bl在不同光照条件下均表现出下胚轴较短的表型。细胞学实验表明,突变体u2bl下胚轴细胞长度的降低是其下胚轴较短的原因。赤霉素(GAs)是促进下胚轴伸长的主效应因子。但u2bl突变体对外源GA处理及内源GA合成抑制剂多效唑(PAC)处理均不敏感,表明U2BL基因可能影响GA的信号转导。亚细胞定位结果表明,U2BL在细胞核中富集。荧光定量Q-PCR分析结果显示,在u2bl突变体中,PRE1、SAUR16、YUC2、YUC8和PIF4等基因的表达均有显著下调,U2BL可能通过调控上述基因来间接调控下胚轴的伸长。本研究结果为进一步探讨U2BL在拟南芥生长发育及在其他物种中可能行使的功能提供了参考。
Abstract:Hypocotyl elongation is essential for early survival and later growth and development in plants. In this investigation, we examined and screened Arabidopsis thaliana (L.) Heynh. mutant phenotypes, identifying the u2bl variant with a notable short hypocotyl. We also conducted preliminary studies on the role of U2BL in the regulation of hypocotyl development in A. thaliana. The u2bl mutant showed a short hypocotyl phenotype under different light conditions. Cytological experiments showed the shorter cell length of hypocotyls in the u2bl mutant was the reason for its short hypocotyl phenotype. The plant hormone gibberellin (GAs) is the main factor promoting hypocotyl elongation, while paclobutrazol (PAC) is an endogenous GA synthesis inhibitor. Our study showed that the u2bl mutant was not sensitive to exogenous GA treatment or PAC treatment, indicating that U2BL affected GA signal transduction. Subcellular localization results indicated that U2BL was enriched in the nucleus. Furthermore, Q-PCR assay showed that the transcription levels of PRE1, SAUR16, YUC2, YUC8, and PIF4 genes were all significantly down-regulated in the u2bl mutant, suggesting that U2BL may indirectly regulate hypocotyl elongation by regulating the above genes. Our study provided a reference for further research on the possible functions of U2BL in the growth and development of A. thaliana and other species.
-
Keywords:
- U2BL gene /
- Arabidopsis thaliana /
- Hypocotyl /
- GA
-
VQ(Valine-glutamine)是植物特有的一类蛋白,因其包含一个特有的保守基序FxxxVQxLTG而得名(F:苯丙氨酸,V:缬氨酸,Q:谷氨酰胺,L:亮氨酸,T:苏氨酸,G:甘氨酸,x 代表任意氨基酸)[1-3]。首个VQ蛋白SIB1(SIGMA FACTOR-BINDING PROTEIN1)在拟南芥(Arabidopsis thaliana (L.) Heynh.)中被发现[4],随后,从拟南芥中又陆续鉴定出33个VQ蛋白。目前,科研人员已在茶(Camellia sinensis (L.) O. Ktze. )[2]、大豆(Glycine max (L.) Merr.)[5]、小麦(Triticum aestivum L.)[6]及烟草(Nicotiana tabacum L.)[3]中分别鉴定出25、75、113和59个VQ基因。
VQ蛋白在植物应答生物与非生物胁迫中发挥着重要作用[7, 8],其作用机制主要有两种。一是通过与WRKY转录因子相互作用发挥功能,如AtVQ23和AtVQ16通过与WRKY33的WRKY结构域结合,增强后者与靶基因的结合能力,从而影响植物的抗病能力[9]。而AtVQ9与WRKY8互作抑制了WRKY8与W-box的结合活性,从而负调控植物的抗盐性[10]。在番茄(Solanum lycopersicum L.)叶片中,SlVQ7与SlWRKY37互作可提高后者的稳定性,并转录激活其下游靶基因,从而正调控茉莉酸及黑暗诱导的叶片衰老[11]。二是作为丝裂原活化蛋白激酶(MAPK)家族的磷酸化底物发挥作用[12-14]。如拟南芥MKS1(AtVQ21)作为AtMPK4的底物,与WRKY25和WRKY33相互作用,有助于激活MPK4调节的病原体防御反应[3]。
多年生黑麦草(Lolium perenne L.)为禾本科早熟禾亚科黑麦草属植物,是世界上广泛种植的冷季型草坪草及牧草[15],具有分蘖多、成坪速度快、粗蛋白含量高、耐牧性好等优点。多年生黑麦草在生长发育过程中常面临高温、盐碱、荫蔽等非生物胁迫,严重影响草坪质量和牧草品质。因此,鉴定多年生黑麦草VQ基因,探究其在应答非生物胁迫中的作用十分必要。然而,目前尚未见相关报道。本研究系统鉴定了多年生黑麦草VQ蛋白家族成员,并对其保守结构域、进化关系、共线性、亚细胞定位及其在高温、盐碱、黑暗等非生物胁迫下的表达模式进行了分析,研究结果旨在为进一步探讨VQ蛋白在多年生黑麦草生长发育及应答非生物逆境胁迫中的作用奠定基础。
1. 材料与方法
1.1 实验材料
本研究以多年生黑麦草品种‘百灵鸟’为实验材料。黑麦草种子于营养土(基质∶蛭石=1∶1)中萌发,萌发后转移至光照培养箱,光周期为16 h / 8 h(光 / 黑暗),温度为25 ℃ / 22 ℃(光 / 黑暗)。待植株生长1个月后进行黑暗、高温和NaCl处理。黑暗处理的条件为:25 ℃ / 22 ℃,24 h黑暗,分别于处理0、2、8 d后取样。高温处理的条件为:42 ℃,光周期16 h / 8 h,分别于处理0、1、24 h后取样。NaCl处理的条件为:25 ℃ / 22 ℃,光周期16 h / 8 h,使用200 mL 200 mmol/L的NaCl溶液进行浇灌,分别于盐处理0、3、38 h后取样。
1.2 多年生黑麦草VQ基因家族成员的鉴定
以Pfam数据库中VQ结构域(PF05678)的隐马尔可夫模型(HMM)图谱作为查询,通过HMMER搜索多年生黑麦草基因组,鉴定了可能的VQ蛋白序列。使用Pfam数据库(http://pfam.xfam.org/)进一步确认了这些序列。在拟南芥基因组数据库(TAIR)中下载拟南芥全基因组序列及其注释信息。
1.3 多年生黑麦草VQ家族成员的理化性质及亚细胞定位
使用在线网站(https://web.expasy.org/compute_pi/)分析多年生黑麦草VQ蛋白的理化性质。使用BIOINF(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-)在线工具对多年生黑麦草VQ家族成员进行亚细胞定位预测。
1.4 多年生黑麦草VQ家族基因系统发育树构建
使用MEGAX(http://www.megasoftware.net)软件的ML法构建系统发育树,进行系统发育分析,并用iTOL(https://itol.embl.de/)网站进行美化。
1.5 多年生黑麦草VQ保守基序及基因结构分析
利用MEME(https://meme-suite.org/meme/tools/meme)网站对VQ蛋白的氨基酸序列进行保守基序分析。结合植物基因组序列和注释文件,用TBtools提取VQ基因的基因组序列及CDS序列,用TBtools软件工具中的Gene Structure View进行可视化。
1.6 VQ基因染色体分布及种内共线性分析
结合植物基因组序列和注释文件,使用TBtools软件工具中的Amazing Gene Location From GFF3/GTF File进行可视化,使用TBtools软件工具中的one step MCScanX进行种内基因组比对,并用Multiple Synreny Plot进行可视化。
1.7 胁迫条件下多年生黑麦草VQ基因的表达模式分析
利用Trizol试剂(Takara,大连)提取多年生黑麦草在不同非生物胁迫处理后的总RNA。使用反转录试剂盒Hifair® Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus)(Yeasen,上海)合成cDNA。利用Hieff® qPCR SYBR Green Master Mix(High Rox Plus)试剂盒(Yeasen,上海)在Step One Plus Real-Time PCR Systems系统上进行qRT-PCR反应。反应体系总体积为20 μL,包括2 μL cDNA,上、下游引物各0.8 μL,10 μL SYBR,用ddH2O补足体积。反应程序为:95 ℃预变性5 min;95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸20 s,40个循环,系统自带熔解曲线。引物序列见附表1
1 ,由擎科公司(擎科,武汉)合成,多年生黑麦草eEF1A(s)为内参基因。采用2−△△Ct方法计算候选基因的相对表达量,实验设3次重复。利用TBtools绘制热图,进行表达模式分析。2. 结果与分析
2.1 LpVQ家族成员鉴定
为了系统分析多年生黑麦草的VQ基因,以VQ结构域(PF05678)为参考,从全基因组水平挖掘其VQ家族基因。结果显示,多年生黑麦草中共存在52个VQ基因。对这些基因的编码产物LpVQ家族成员进行理化性质分析,发现LpVQs的氨基酸长度介于118~427 aa,蛋白分子量范围为11.67~43.84 kD,等电点在5.01~11.6。通过BIOINF在线网站对LpVQ家族成员进行亚细胞定位预测,发现48个蛋白定位在细胞核中,LpVQ22为叶绿体与细胞核共定位,LpVQ33为细胞壁与细胞核共定位,LpVQ37为细胞膜与细胞核共定位,而LpVQ45预测定位在细胞膜中(附表2
1 )。2.2 LpVQ家族系统进化树分析
为了分析LpVQs的进化关系,利用多年生黑麦草的52个LpVQ蛋白和拟南芥的34个AtVQ蛋白共同构建系统进化树。结果显示,拟南芥与多年生黑麦草的VQ家族成员可划分为7个亚家族(Ⅰ~Ⅶ),分别包含5、8、6、11、12、6和4个LpVQ蛋白。亚家族Ⅰ中只包含LpVQs,亚家族Ⅳ、Ⅴ和Ⅵ中含有较多的LpVQ家族成员,而AtVQ家族成员则更多地归类于亚家族Ⅱ、Ⅲ和Ⅶ(图1)。进一步分析发现,多年生黑麦草与拟南芥的VQ家族间存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。
![]() 图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne
图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne2.3 LpVQ家族成员保守基序和基因结构分析
为了研究蛋白的结构多样性,利用MEME在线软件对52个LpVQ蛋白的保守基序进行分析,结果发现17%的LpVQs含有1个Motif,48%的LpVQs含有两个Motif,19%的LpVQs含有3个Motif,6%的LpVQs含有4个Motif,10%的LpVQs含有5个Motif。所有家族成员均具有Motif1,表明它是LpVQ家族的核心保守结构域(图2)。
此外,本研究用TBtools工具对LpVQs进行了基因结构分析。结果发现,46个LpVQ基因没有内含子,5个LpVQ基因含有1个内含子(LpVQ24、LpVQ29、LpVQ31、LpVQ32、LpVQ52),仅有1个基因(LpVQ161)含有两个内含子(附图1
1 )。2.4 LpVQ家族成员染色体定位以及共线性分析
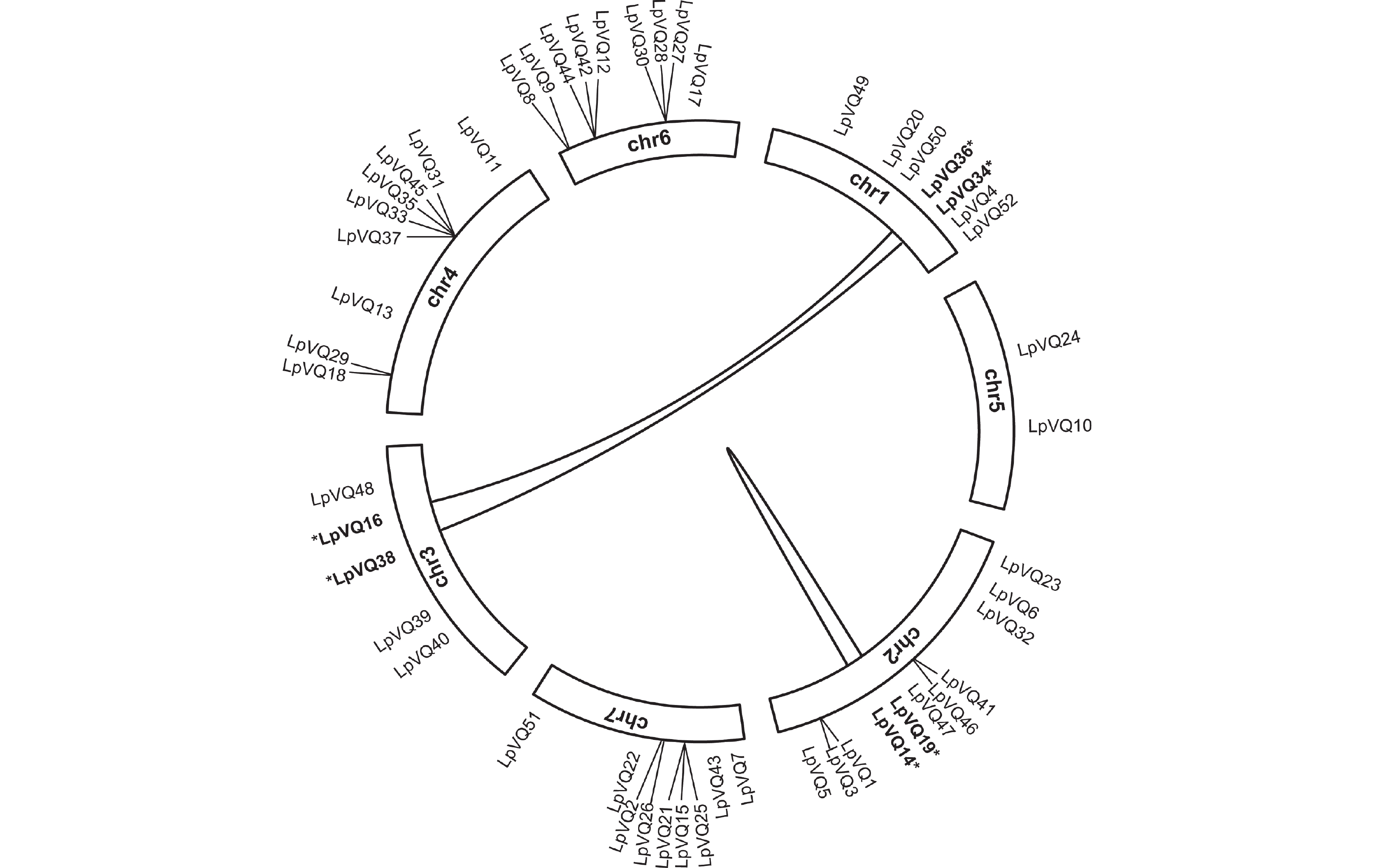
染色体定位分析结果表明,LpVQ基因分布在多年生黑麦草的7条染色体上,其中分布在chr1上的有7个,占13%,分布在chr2上的最多,为11个,占21%,分布在chr3上的为5个,占10%,分布在chr4、chr6和chr7上的基因均各有9个,各占17%,而分布在chr5上的最少,仅有2个,占4%(图3)。
为了了解LpVQ基因之间的进化关系,本研究对VQ家族成员进行了种内共线性分析。结果表明,3对VQ基因之间存在共同进化关系,分别是LpVQ16与LpVQ36、LpVQ34与LpVQ38以及LpVQ14与LpVQ19(图4)。
2.5 LpVQ基因响应非生物胁迫的表达模式分析
从7个亚家族中分别随机选取3个LpVQ基因,检测其在黑暗、高温以及盐胁迫下多年生黑麦草中的表达量变化。结果显示,黑暗处理后,只有4个LpVQ基因的表达持续上调。大部分LpVQ基因的表达量在黑暗处理2 d后下降,其中,LpVQ2、LpVQ8、LpVQ14、LpVQ22及LpVQ38的表达量在黑暗处理8 d后又显著上升(图5:A)。
![]() 图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.
图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.42 ℃高温处理后,LpVQ的表达模式被分为3类。LpVQ18、LpVQ8、LpVQ28、LpVQ2、LpVQ7、LpVQ4及LpVQ12受1 h高温处理的显著诱导,而高温处理24 h后,其表达又显著下调。LpVQ9、LpVQ22、LpVQ15、LpVQ3、LpVQ42、LpVQ6、LpVQ10、LpVQ1、LpVQ38的表达量在1 h和24 h高温处理后均显著下降。LpVQ5、LpVQ27、LpVQ34、LpVQ14和LpVQ44的表达量只在24 h高温处理后下降(图5:B)。
NaCl处理3 h后,13个LpVQ基因显著上调表达,其中LpVQ4的表达上升了13倍。而NaCl处理38 h后,大部分基因的表达明显下调。仅有3个基因的表达受盐诱导持续上调,分别是 LpVQ8、LpVQ10和LpVQ38(图5:C)。
3. 讨论
近年来,VQ基因已经在多种植物中被发掘鉴定,包括拟南芥、水稻(Oryza sativa L.)、大豆和葡萄(Vitis vinifera L.)等[16-19],而关于多年生黑麦草VQ家族基因的鉴定和分析还鲜见报道。本研究通过全基因组鉴定,得到52个多年生黑麦草LpVQ基因,并系统分析了其理化特性、蛋白结构、基因结构、共线性、进化关系以及响应逆境胁迫的表达模式。
基因结构分析发现,88.46%的LpVQ基因无内含子,与之类似,在拟南芥[16]、水稻[17]、玉米(Zea mays L.)[20]和马铃薯(Solanum tuberosum L.)[21]中,无内含子的VQ基因分别占88.2%、92.5%、88.5%和91.89%。而苔藓(Bryophyta)[22]中无内含子的VQ基因比例则远低于以上物种,仅为28.0%,说明VQ基因可能在长期进化过程中丢失了内含子。
系统进化分析结果显示,多年生黑麦草和拟南芥的VQ家族成员可被分为7个亚家族,且LpVQs与AtVQs亲缘关系较远,只存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。根据拟南芥VQ基因的功能,可以推测多年生黑麦草中其直系同源VQ基因的功能。有研究指出,AtVQ9作为AtWRKY8的抑制子,能提高拟南芥的耐盐性[9],由此推测,其直系同源基因LpVQ11可能也在多年生黑麦草抵御盐胁迫中发挥重要作用。
VQ基因在植物响应非生物胁迫中发挥着重要作用。如,过表达PtVQ1基因显著增强了转基因拟南芥的耐盐性[23];而过表达MdVQ37则提高了转基因苹果(Malus pumila Mill.)对高温胁迫的敏感性[24]。本研究对黑暗、高温及盐胁迫处理下多年生黑麦草的VQ表达量进行了分析。结果显示,LpVQ基因广泛响应黑暗、高温及盐胁迫,且大部分LpVQ基因均在胁迫初期即做出快速响应,这与西葫芦(Cucurbita pepo L.)[8]、黄瓜(Cucumis sativus L.)[25]等物种中的研究结果相类似。然而,LpVQ基因的表达模式在不同胁迫处理下也存在差异。如,LpVQ1在黑暗胁迫处理后持续下调表达,而在高温和盐胁迫下持续上调表达,推测该基因可能在不同胁迫中发挥着不同的作用。尽管LpVQ可能参与多年生黑麦草对黑暗、高温及盐等胁迫响应的调控,但其具体功能还需要进一步验证。
综上所述,本研究对多年生黑麦草VQ基因家族成员进行了系统鉴定与分析,为后续验证VQ家族成员在多年生黑麦草应答生物或非生物胁迫中的功能提供了基础。
-
![]()
图 1 u2bl突变体的获得与鉴定
A:U2BL的基因结构示意图(黑框代表外显子,直线部分代表内含子,白色倒三角代表T-DNA片段插入位点);B:u2bl突变体的基因型鉴定;C:u2bl突变体的半定量RT-PCR电泳图。
Figure 1. Acquisition and genotyping of u2bl mutants
A: Gene structure schematic of U2BL (Black boxes represent exons, black line segments represent introns, and inverted triangles represent T-DNA fragment insertion sites); B: Genotype identification of u2bl mutants; C: RT-PCR result of u2bl mutants.
![]()
图 2 u2bl突变体及野生型下胚轴表型观察
A:长日照(16 h光照/8 h黑暗)下u2bl突变体的下胚轴表型图;B:短日照(8 h光照/16 h黑暗)下u2bl突变体的下胚轴表型图;C~E:长日照、短日照以及黑暗条件下的下胚轴长度统计图。统计40株,数据表示为平均值 ± 标准误差,实验重复3次。标尺为0.5 mm。* 表示P<0.05,下同。
Figure 2. Phenotypic observation of hypocotyl in u2bl mutants and wild-type
A: Phenotype of hypocotyl of u2bl mutants under long-day conditions (16 h light/8 h dark); B: Hypocotyl phenotype of u2bl mutants under short-day conditions (8h light/16 h dark); C-E: Statistical chart of hypocotyl length under long-day, short-day, and dark conditions. There were 40 samples, and data is represented as mean ± standard error (SE). Experiment was repeated independently three times. Scale is 0.5 mm. * indicates P<0.05. Same below.
![]()
图 3 u2bl突变体及野生型的下胚轴细胞数目与长度
A:长日照条件下培养7 d的下胚轴细胞长度观察,白线指示细胞大小;B、C:长日照条件下培养7 d的下胚轴细胞个数和细胞长度统计值,B图是幼苗下胚轴一列细胞的个数,C图是细胞下胚轴长度/幼苗下胚轴一列细胞个数(n≥30)。标尺为0.1 µm。ns: 在0.05水平上差异不显著。
Figure 3. Cell numbers and lengths in hypocotyl of u2bl mutants and wild-type
A: Hypocotyls of Col-0, u2bl-1, and u2bl-2 cultured for 7 d under long-day conditions; B, C: Number and length of hypocotyl cells of Col-0, u2bl-1, and u2bl-2 cultured for 7 d under long-day conditions (n≥30). Fig. 3: B shows the number of cells in a row of seedling hypocotyls, Fig. 3: C is the length of cell hypocotyls/number of cells in a row of seedling hypocotyls. Scale is 0.1 µm. ns: Not significant at P<0.05 level.
![]()
图 4 u2bl突变体和野生型对GA和PAC的敏感性
A:长日照条件下,不同浓度GA对突变体及野生型下胚轴长度的影响;B:根据A图结果进行的相对增长率统计;C:长日照条件下,不同浓度PAC对突变体及野生型下胚轴长度影响;D:根据C图结果进行的相对增长率统计。
Figure 4. Sensitivity of u2bl mutant and wild-type to GA and PAC
A: Under long-day conditions, the effect of different concentrations of GA on the length of hypocotyls in mutant and wild-type plants; B: According to the results in Fig. A, relative growth rates of Col-0, u2bl-1, u2bl-2, and rga gai rgl1 were calculated; C: Under long-day conditions, the effect of different concentrations of PAC on the length of hypocotyls in mutant and wild-type plants; D: According to the results in Fig. C, relative growth rate of Col-0, u2bl-1, u2bl-2, and rga gai rgl1 was calculated.
![]()
图 5 U2BL的亚细胞定位
A:过表达U2BL转基因植株根尖细胞荧光信号定位。B:拟南芥原生质体中,红色为U2BL-RFP融合的信号,蓝色为DAPI。BF为明场。
Figure 5. Subcellular localization of U2BL
A: Transgenic plants overexpressed U2BL were grown in medium for 7 d, and fluorescence signal localization of root tip cells was observed by laser confocal microscopy. B: In Arabidopsis thaliana protoplast, red is signal of U2BL-RFP fusion, and blue is DAPI. BF is brightfield.
![]()
图 6 Q-PCR检测GA信号通路下游基因的表达
Figure 6. Partial downstream gene expression in GA signaling pathway by Q-PCR
表 1 实验所用引物
Table 1 Primers used in this study
引物
Primers序列(5'–3')
Sequence (5'–3')LP1 AGTTCTGCATCTGACGGACAG RP1 CGGCGAGATAGACACTCAGAC LP2 AAAAGGGGAAAGAGATAGGGC RP2 CCACAGAGATCACACATCACG LBP1.3 ATTTTGCCGATTTCGGAAC TUBULIN CTTAAGCTCACCACTCCAAGCT TUBULIN GCACTTCCACTTCGTCTTCTTC PRE1-F GTTCTGATAAGGCATCAGCCTCG PRE1-R CATGAGTAGGCTTCTAATAACGG PRE5-F AACGGCGTCGTTCTGATAAG PRE5-R CATGAGTAAGCTTCTAATCACGG PRE6-F TCCAACACCTCATCCCTGAACTTC PRE6-R CGGTCACTGAGGTCATCAACCTCTC YUC2-F ATCCGAAGTGGACACATCAA YUC2-R TTAGCCACATGGGTACGTTG YUC8-F GACAGGATTGTATGCGGTTG YUC8-R TTCAAGTTGCCAAACAGAGC PIF4-F CGACCGGTTTGCTAGATACA PIF4-R GACGACGGTTGTTGACTTTG SAUR16-F AAATCTCGGGTTTTAGTTCGTCA SAUR16-R TGCAAGCCACCACTTATCAGC UBQ10 AGGATGGCAGAACTCTTGCT UBQ10 TCCCAGTCAACGTCTTAACG SAUR19-F ACGTCGTCTCAAGCAGCATCTATCA SAUR19-R CCCACGTAAACCGGAAAATGACCTT  下载: 导出CSV
下载: 导出CSV
-
[1] Boron AK,Vissenberg K. The Arabidopsis thaliana hypocotyl,a model to identify and study control mechanisms of cellular expansion[J]. Plant Cell Rep,2014,33(5):697−706. doi: 10.1007/s00299-014-1591-x
[2] Quail PH. Phytochrome photosensory signalling networks[J]. Nat Rev Mol Cell Biol,2002,3(2):85−93.
[3] Rizzini L,Favory JJ,Cloix C,Faggionato D,O’hara A,et al. Perception of UV-B by the Arabidopsis UVR8 protein[J]. Science,2011,332(6025):103−106. doi: 10.1126/science.1200660
[4] McNellis TW,Deng XW. Light control of seedling morphogenetic pattern[J]. Plant Cell,1995,7(11):1749−1761.
[5] Deng XW,Quail PH. Signalling in light-controlled development[J]. Semin Cell Dev Biol,1999,10(2):121−129. doi: 10.1006/scdb.1999.0287
[6] Lian HL,He SB,Zhang YC,Zhu DM,Zhang JY,et al. Blue-light-dependent interaction of cryptochrome 1 with SPA1 defines a dynamic signaling mechanism[J]. Genes Dev,2011,25(10):1023−1028. doi: 10.1101/gad.2025111
[7] Liu B,Zuo ZC,Liu HT,Liu XM,Lin CT. Arabidopsis cryptochrome 1 interacts with SPA1 to suppress COP1 activity in response to blue light[J]. Genes Dev,2011,25(10):1029−1034. doi: 10.1101/gad.2025011
[8] Zuo ZC,Liu HT,Liu B,Liu XM,Lin CT. Blue light-dependent interaction of CRY2 with SPA1 regulates COP1 activity and floral initiation in Arabidopsis[J]. Curr Biol,2011,21(10):841−847. doi: 10.1016/j.cub.2011.03.048
[9] Quail PH,Boylan MT,Parks BM,Short TW,Xu Y,Wagner D. Phytochromes:photosensory perception and signal transduction[J]. Science,1995,268(5211):675−680. doi: 10.1126/science.7732376
[10] Somers DE,Sharrock RA,Tepperman JM,Quail PH. The hy3 long hypocotyl mutant of Arabidopsis is deficient in phytochrome B[J]. Plant Cell,1991,3(12):1263−1274. doi: 10.2307/3869307
[11] Huq E,Al-Sady B,Hudson M,Kim C,Apel K,Quail PH. Phytochrome-interacting factor 1 is a critical bHLH regulator of chlorophyll biosynthesis[J]. Science,2004,305(5692):1937−1941. doi: 10.1126/science.1099728
[12] Leivar P,Quail PH. PIFs:pivotal components in a cellular signaling hub[J]. Trends Plant Sci,2011,16(1):19−28. doi: 10.1016/j.tplants.2010.08.003
[13] Al-Sady B,Ni WM,Kircher S,Schäfer E,Quail PH. Photoactivated phytochrome induces rapid PIF3 phosphorylation prior to proteasome-mediated degradation[J]. Mol Cell,2006,23(3):439−446. doi: 10.1016/j.molcel.2006.06.011
[14] Shen Y,Khanna R,Carle CM,Quail PH. Phytochrome induces rapid PIF5 phosphorylation and degradation in response to red-light activation[J]. Plant Physiol,2007,145(3):1043−1051. doi: 10.1104/pp.107.105601
[15] Shen H,Zhu L,Castillon A,Majee M,Downie B,Huq E. Light-induced phosphorylation and degradation of the negative regulator PHYTOCHROME-INTERACTING FACTOR1 from Arabidopsis depend upon its direct physical interactions with photoactivated phytochromes[J]. Plant Cell,2008,20(6):1586−1602. doi: 10.1105/tpc.108.060020
[16] Quint M,Delker C,Franklin KA,Wigge PA,Halliday KJ,van Zanten M. Molecular and genetic control of plant thermomorphogenesis[J]. Nat Plants,2016,2(1):15190. doi: 10.1038/nplants.2015.190
[17] Casal JJ,Balasubramanian S. Thermomorphogenesis[J]. Annu Rev Plant Biol,2019,70:321−346. doi: 10.1146/annurev-arplant-050718-095919
[18] Gil KE,Park CM. Thermal adaptation and plasticity of the plant circadian clock[J]. New Phytol,2019,221(3):1215−1229. doi: 10.1111/nph.15518
[19] Fiorucci AS,Galvão VC,Ince YÇ,Boccaccini A,Goyal A,et al. PHYTOCHROME INTERACTING FACTOR 7 is important for early responses to elevated temperature in Arabidopsis seedlings[J]. New Phytol,2020,226(1):50−58. doi: 10.1111/nph.16316
[20] Bai MY,Shang JX,Oh E,Fan M,Bai Y,et al. Brassinosteroid,gibberellin and phytochrome impinge on a common transcription module in Arabidopsis[J]. Nat Cell Biol,2012,14(8):810−817. doi: 10.1038/ncb2546
[21] 黄炎霞. 赤霉素调控的南方水稻黑条矮缩病的发病[D]. 福州: 福建农林大学, 2014: 106. [22] Hedden P,Sponsel V. A century of gibberellin research[J]. J Plant Growth Regul,2015,34(4):740−760. doi: 10.1007/s00344-015-9546-1
[23] Takehara S,Sakuraba S,Mikami B,Yoshida H,Yoshimura H,et al. A common allosteric mechanism regulates homeostatic inactivation of auxin and gibberellin[J]. Nat Commun,2020,11(1):2143. doi: 10.1038/s41467-020-16068-0
[24] 李欣. 大豆GmGAMYB基因调控花期与株高的功能研究[D]. 哈尔滨: 东北农业大学, 2021: 68. [25] 李保珠,赵翔,安国勇. 赤霉素的研究进展[J]. 中国农学通报,2011,27(1):1−5. Li BZ,Zhao X,An GY. Recent advances in research of gibberellin[J]. Chinese Agricultural Science Bulletin,2011,27(1):1−5.
[26] De Lucas M,Davière JM,Rodríguez-Falcón M,Pontin M,Iglesias-Pedraz JM,et al. A molecular framework for light and gibberellin control of cell elongation[J]. Nature,2008,451(7177):480−484. doi: 10.1038/nature06520
[27] Feng SH,Martinez C,Gusmaroli G,Wang Y,Zhou JL,et al. Coordinated regulation of Arabidopsis thaliana development by light and gibberellins[J]. Nature,2008,451(7177):475−479. doi: 10.1038/nature06448
[28] Ueguchi-Tanaka M,Ashikari M,Nakajima M,Itoh H,Katoh E,et al. GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin[J]. Nature,2005,437(7059):693−698. doi: 10.1038/nature04028
[29] Nakajima M,Shimada A,Takashi Y,Kim YC,Park SH,et al. Identification and characterization of Arabidopsis gibberellin receptors[J]. Plant J,2006,46(5):880−889. doi: 10.1111/j.1365-313X.2006.02748.x
[30] Griffiths J,Murase K,Rieu I,Zentella R,Zhang ZL,et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis[J]. Plant Cell,2006,18(12):3399−3414.
[31] Willige BC,Ghosh S,Nill C,Zourelidou M,Dohmann EMN,et al. The DELLA domain of GA INSENSITIVE mediates the interaction with the GA INSENSITIVE DWARF1A gibberellin receptor of Arabidopsis[J]. Plant Cell,2007,19(4):1209−1220. doi: 10.1105/tpc.107.051441
[32] Suzuki H,Park SH,Okubo K,Kitamura J,Ueguchi-tanaka M,et al. Differential expression and affinities of Arabidopsis gibberellin receptors can explain variation in phenotypes of multiple knock-out mutants[J]. Plant J,2009,60(1):48−55. doi: 10.1111/j.1365-313X.2009.03936.x
[33] Cheng H,Qin LJ,Lee S,Fu XD,Richards DE,et al. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function[J]. Development,2004,131(5):1055−1064. doi: 10.1242/dev.00992
[34] Jangi M,Sharp PA. Building robust transcriptomes with master splicing factors[J]. Cell,2014,159(3):487−498. doi: 10.1016/j.cell.2014.09.054
[35] Vandivier LE,Anderson SJ,Foley SW,Gregory BD. The conservation and function of RNA secondary structure in plants[J]. Annu Rev Plant Biol,2016,67:463−488. doi: 10.1146/annurev-arplant-043015-111754
[36] Cho H,Cho HS,Hwang I. Emerging roles of RNA-binding proteins in plant development[J]. Curr Opin Plant Biol,2019,51:51−57. doi: 10.1016/j.pbi.2019.03.016
[37] Staněk D,Neugebauer KM. The Cajal body:a meeting place for spliceosomal snRNPs in the nuclear maze[J]. Chromosoma,2006,115(5):343−354. doi: 10.1007/s00412-006-0056-6
[38] Calixto CPG,Guo WB,James AB,Tzioutziou NA,Entizne JC,et al. Rapid and dynamic alternative splicing impacts the Arabidopsis cold response transcriptome[J]. Plant Cell,2018,30(7):1424−1444. doi: 10.1105/tpc.18.00177
[39] Fukazawa J, Ito T, Kamiya Y, Yamaguchi S, Takahashi Y. Binding of GID1 to DELLAs promotes dissociation of GAF1 from DELLA in GA dependent manner. Plant Signal Behav, 2015, 10(10): e1052923.
[40] Ito T, Okada K, Fukazawa J, Takahashi Y. DELLA-dependent and -independent gibberellin signaling. Plant Signal Behav, 2018, 13(3): e1445933.










计量
- 文章访问数: 342
- HTML全文浏览量: 28
- PDF下载量: 125
