
Interspecific relationship between Armeniaca hongpingensis T. T. Yu & C. L. Li and related species based on morphological characters
-
摘要:
本研究通过野外调查获得洪平杏(Armeniaca hongpingensis T. T. Yu & C. L. Li)、杏(A. vulgaris Lam.)、梅(A. mume Siebold)和杏梅( A. mume var. bungo Makino)有关的叶、花、果、果核及枝条的21个定量性状和15个定性性状,并对这些数据进行了聚类分析。结果显示,无论是PAM聚类还是UPGMA聚类分析都可清晰地将4种杏属植物分开,说明物种之间具有明显的区分特征。主成分分析的前3个主成分中,一年生枝颜色、被毛情况,叶及果实被毛,叶形指标(叶形、叶尖、叶基)和果核表面特征的绝对权重值都在0.9以上,表明这些性状在4种杏属植物的分类中具有重要的作用。电镜扫描观察结果发现,供试花粉均是单粒花粉,等极,辐射对称,具3孔萌发沟。杏、梅、杏梅的花粉表面纹饰均为条纹状,但洪平杏的表面纹饰为间断条纹,且有不明显穿孔。4种植物叶上表皮细胞均具条纹状角质增厚。与其他种相比,洪平杏细胞表面条纹常弯曲、交叠,无固定走向。根据聚类分析结果和微形态特征,支持将洪平杏作为区别于梅和杏的一个独立种的处理。
Abstract:Armeniaca hongpingensis T. T. Yu & C. L. Li is a critically endangered species distributed only in Hongping Town, Shennongjia, Hubei Province, China. According to the Flora of China, A. hongpingensis and A. mume var. bungo Makino may be natural hybrids of A. mume Lam. and A. vulgaris Siebold. To date, however, no morphological study on the relationship between A. hongpingensis, A. vulgaris, A. mume, and A. mume var. bungo has been conducted. Through field investigations, 21 quantitative and 15 qualitative leaf, flower, fruit, kernel, and branch characters were obtained, followed by cluster analysis. Results showed that both PAM and UPGMA cluster analysis clearly divided the specimens into four species, indicating obvious distinguishing characters among them. Based on principal component analysis, the absolute weight values of annual branch color and hair-covering status, hair-covering status of leaf and fruit, leaf shape index (leaf shape, leaf tip, leaf base), and surface characters of kernel in the first three principal components were all above 0.9, indicating that these traits play important roles in the classification of the four Armeniaca species. Based on electron microscopy, the pollen grains were identified as single grain (monads), isopolar, radially symmetrical, and tri-colporate. The pollen surfaces of A. vulgaris, A. mume, and A. mume var. bungo were striate-ornamented, while the pollen of A. hongpingensis was discontinuously striped and showed inconspicuous perforation. The epidermal cells of the leaves of the four plants all showed striate keratinization. Compared to the other species, the stripes on the cell surface of A. hongpingensis were often curved, overlapped, and without fixed direction. According to cluster analysis and micromorphological characters, A. hongpingensis should be treated as an independent species from A. vulgaris and A. mume.
-
Keywords:
- Armeniaca hongpingensis /
- Interspecific relationship /
- Micromorphology /
- Palynology
-
VQ(Valine-glutamine)是植物特有的一类蛋白,因其包含一个特有的保守基序FxxxVQxLTG而得名(F:苯丙氨酸,V:缬氨酸,Q:谷氨酰胺,L:亮氨酸,T:苏氨酸,G:甘氨酸,x 代表任意氨基酸)[1-3]。首个VQ蛋白SIB1(SIGMA FACTOR-BINDING PROTEIN1)在拟南芥(Arabidopsis thaliana (L.) Heynh.)中被发现[4],随后,从拟南芥中又陆续鉴定出33个VQ蛋白。目前,科研人员已在茶(Camellia sinensis (L.) O. Ktze. )[2]、大豆(Glycine max (L.) Merr.)[5]、小麦(Triticum aestivum L.)[6]及烟草(Nicotiana tabacum L.)[3]中分别鉴定出25、75、113和59个VQ基因。
VQ蛋白在植物应答生物与非生物胁迫中发挥着重要作用[7, 8],其作用机制主要有两种。一是通过与WRKY转录因子相互作用发挥功能,如AtVQ23和AtVQ16通过与WRKY33的WRKY结构域结合,增强后者与靶基因的结合能力,从而影响植物的抗病能力[9]。而AtVQ9与WRKY8互作抑制了WRKY8与W-box的结合活性,从而负调控植物的抗盐性[10]。在番茄(Solanum lycopersicum L.)叶片中,SlVQ7与SlWRKY37互作可提高后者的稳定性,并转录激活其下游靶基因,从而正调控茉莉酸及黑暗诱导的叶片衰老[11]。二是作为丝裂原活化蛋白激酶(MAPK)家族的磷酸化底物发挥作用[12-14]。如拟南芥MKS1(AtVQ21)作为AtMPK4的底物,与WRKY25和WRKY33相互作用,有助于激活MPK4调节的病原体防御反应[3]。
多年生黑麦草(Lolium perenne L.)为禾本科早熟禾亚科黑麦草属植物,是世界上广泛种植的冷季型草坪草及牧草[15],具有分蘖多、成坪速度快、粗蛋白含量高、耐牧性好等优点。多年生黑麦草在生长发育过程中常面临高温、盐碱、荫蔽等非生物胁迫,严重影响草坪质量和牧草品质。因此,鉴定多年生黑麦草VQ基因,探究其在应答非生物胁迫中的作用十分必要。然而,目前尚未见相关报道。本研究系统鉴定了多年生黑麦草VQ蛋白家族成员,并对其保守结构域、进化关系、共线性、亚细胞定位及其在高温、盐碱、黑暗等非生物胁迫下的表达模式进行了分析,研究结果旨在为进一步探讨VQ蛋白在多年生黑麦草生长发育及应答非生物逆境胁迫中的作用奠定基础。
1. 材料与方法
1.1 实验材料
本研究以多年生黑麦草品种‘百灵鸟’为实验材料。黑麦草种子于营养土(基质∶蛭石=1∶1)中萌发,萌发后转移至光照培养箱,光周期为16 h / 8 h(光 / 黑暗),温度为25 ℃ / 22 ℃(光 / 黑暗)。待植株生长1个月后进行黑暗、高温和NaCl处理。黑暗处理的条件为:25 ℃ / 22 ℃,24 h黑暗,分别于处理0、2、8 d后取样。高温处理的条件为:42 ℃,光周期16 h / 8 h,分别于处理0、1、24 h后取样。NaCl处理的条件为:25 ℃ / 22 ℃,光周期16 h / 8 h,使用200 mL 200 mmol/L的NaCl溶液进行浇灌,分别于盐处理0、3、38 h后取样。
1.2 多年生黑麦草VQ基因家族成员的鉴定
以Pfam数据库中VQ结构域(PF05678)的隐马尔可夫模型(HMM)图谱作为查询,通过HMMER搜索多年生黑麦草基因组,鉴定了可能的VQ蛋白序列。使用Pfam数据库(http://pfam.xfam.org/)进一步确认了这些序列。在拟南芥基因组数据库(TAIR)中下载拟南芥全基因组序列及其注释信息。
1.3 多年生黑麦草VQ家族成员的理化性质及亚细胞定位
使用在线网站(https://web.expasy.org/compute_pi/)分析多年生黑麦草VQ蛋白的理化性质。使用BIOINF(http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-)在线工具对多年生黑麦草VQ家族成员进行亚细胞定位预测。
1.4 多年生黑麦草VQ家族基因系统发育树构建
使用MEGAX(http://www.megasoftware.net)软件的ML法构建系统发育树,进行系统发育分析,并用iTOL(https://itol.embl.de/)网站进行美化。
1.5 多年生黑麦草VQ保守基序及基因结构分析
利用MEME(https://meme-suite.org/meme/tools/meme)网站对VQ蛋白的氨基酸序列进行保守基序分析。结合植物基因组序列和注释文件,用TBtools提取VQ基因的基因组序列及CDS序列,用TBtools软件工具中的Gene Structure View进行可视化。
1.6 VQ基因染色体分布及种内共线性分析
结合植物基因组序列和注释文件,使用TBtools软件工具中的Amazing Gene Location From GFF3/GTF File进行可视化,使用TBtools软件工具中的one step MCScanX进行种内基因组比对,并用Multiple Synreny Plot进行可视化。
1.7 胁迫条件下多年生黑麦草VQ基因的表达模式分析
利用Trizol试剂(Takara,大连)提取多年生黑麦草在不同非生物胁迫处理后的总RNA。使用反转录试剂盒Hifair® Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR(gDNA digester plus)(Yeasen,上海)合成cDNA。利用Hieff® qPCR SYBR Green Master Mix(High Rox Plus)试剂盒(Yeasen,上海)在Step One Plus Real-Time PCR Systems系统上进行qRT-PCR反应。反应体系总体积为20 μL,包括2 μL cDNA,上、下游引物各0.8 μL,10 μL SYBR,用ddH2O补足体积。反应程序为:95 ℃预变性5 min;95 ℃变性10 s,60 ℃退火20 s,72 ℃延伸20 s,40个循环,系统自带熔解曲线。引物序列见附表1
1 ,由擎科公司(擎科,武汉)合成,多年生黑麦草eEF1A(s)为内参基因。采用2−△△Ct方法计算候选基因的相对表达量,实验设3次重复。利用TBtools绘制热图,进行表达模式分析。2. 结果与分析
2.1 LpVQ家族成员鉴定
为了系统分析多年生黑麦草的VQ基因,以VQ结构域(PF05678)为参考,从全基因组水平挖掘其VQ家族基因。结果显示,多年生黑麦草中共存在52个VQ基因。对这些基因的编码产物LpVQ家族成员进行理化性质分析,发现LpVQs的氨基酸长度介于118~427 aa,蛋白分子量范围为11.67~43.84 kD,等电点在5.01~11.6。通过BIOINF在线网站对LpVQ家族成员进行亚细胞定位预测,发现48个蛋白定位在细胞核中,LpVQ22为叶绿体与细胞核共定位,LpVQ33为细胞壁与细胞核共定位,LpVQ37为细胞膜与细胞核共定位,而LpVQ45预测定位在细胞膜中(附表2
1 )。2.2 LpVQ家族系统进化树分析
为了分析LpVQs的进化关系,利用多年生黑麦草的52个LpVQ蛋白和拟南芥的34个AtVQ蛋白共同构建系统进化树。结果显示,拟南芥与多年生黑麦草的VQ家族成员可划分为7个亚家族(Ⅰ~Ⅶ),分别包含5、8、6、11、12、6和4个LpVQ蛋白。亚家族Ⅰ中只包含LpVQs,亚家族Ⅳ、Ⅴ和Ⅵ中含有较多的LpVQ家族成员,而AtVQ家族成员则更多地归类于亚家族Ⅱ、Ⅲ和Ⅶ(图1)。进一步分析发现,多年生黑麦草与拟南芥的VQ家族间存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。
![]() 图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne
图 1 拟南芥和多年生黑麦草VQ家族成员系统进化树分析Figure 1. Phylogenetic analysis of VQ family members in Arabidopsis thaliana and Lolium perenne2.3 LpVQ家族成员保守基序和基因结构分析
为了研究蛋白的结构多样性,利用MEME在线软件对52个LpVQ蛋白的保守基序进行分析,结果发现17%的LpVQs含有1个Motif,48%的LpVQs含有两个Motif,19%的LpVQs含有3个Motif,6%的LpVQs含有4个Motif,10%的LpVQs含有5个Motif。所有家族成员均具有Motif1,表明它是LpVQ家族的核心保守结构域(图2)。
此外,本研究用TBtools工具对LpVQs进行了基因结构分析。结果发现,46个LpVQ基因没有内含子,5个LpVQ基因含有1个内含子(LpVQ24、LpVQ29、LpVQ31、LpVQ32、LpVQ52),仅有1个基因(LpVQ161)含有两个内含子(附图1
1 )。2.4 LpVQ家族成员染色体定位以及共线性分析
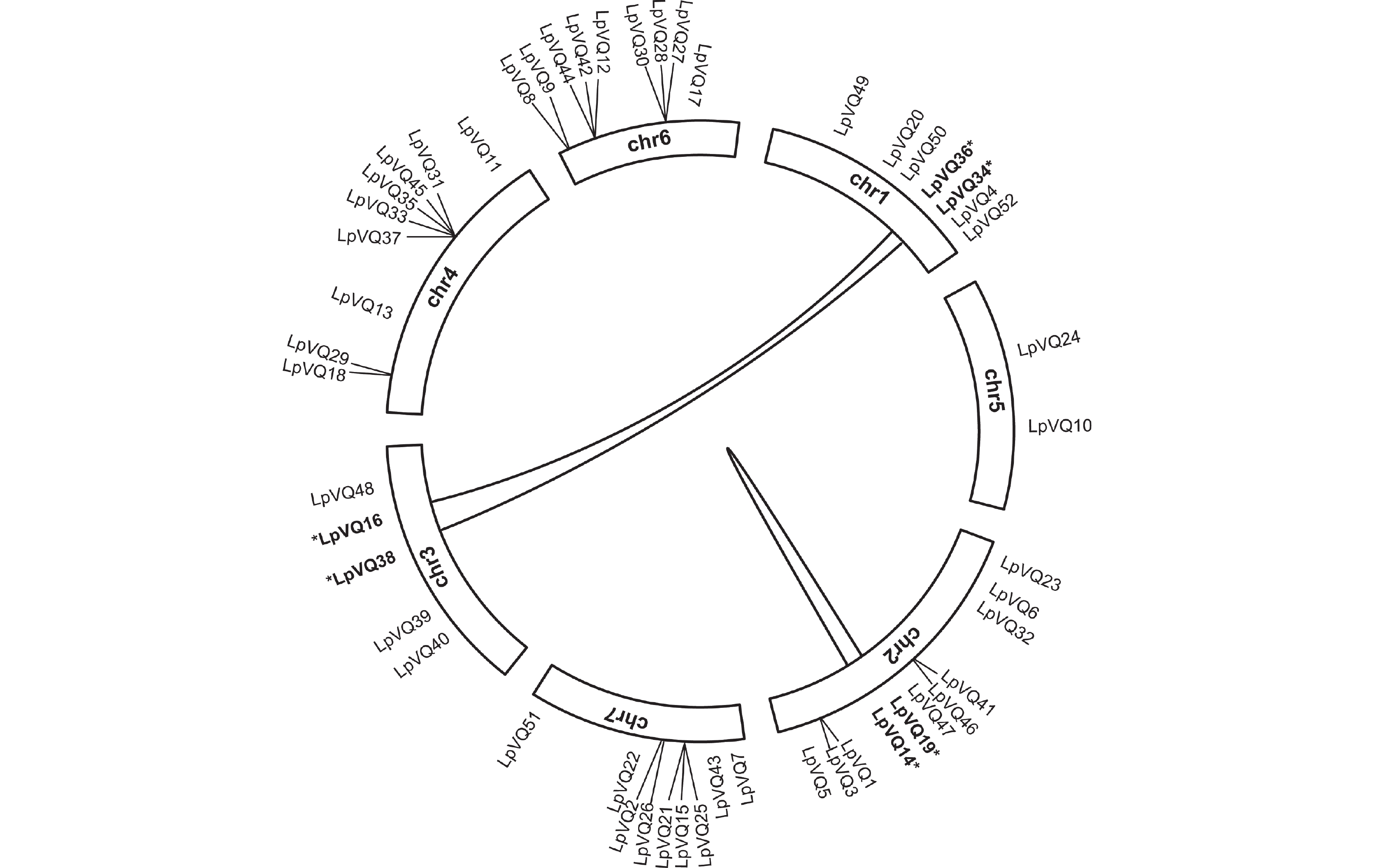
染色体定位分析结果表明,LpVQ基因分布在多年生黑麦草的7条染色体上,其中分布在chr1上的有7个,占13%,分布在chr2上的最多,为11个,占21%,分布在chr3上的为5个,占10%,分布在chr4、chr6和chr7上的基因均各有9个,各占17%,而分布在chr5上的最少,仅有2个,占4%(图3)。
为了了解LpVQ基因之间的进化关系,本研究对VQ家族成员进行了种内共线性分析。结果表明,3对VQ基因之间存在共同进化关系,分别是LpVQ16与LpVQ36、LpVQ34与LpVQ38以及LpVQ14与LpVQ19(图4)。
2.5 LpVQ基因响应非生物胁迫的表达模式分析
从7个亚家族中分别随机选取3个LpVQ基因,检测其在黑暗、高温以及盐胁迫下多年生黑麦草中的表达量变化。结果显示,黑暗处理后,只有4个LpVQ基因的表达持续上调。大部分LpVQ基因的表达量在黑暗处理2 d后下降,其中,LpVQ2、LpVQ8、LpVQ14、LpVQ22及LpVQ38的表达量在黑暗处理8 d后又显著上升(图5:A)。
![]() 图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.
图 5 LpVQ基因响应黑暗、高温及NaCl胁迫的表达模式分析A:黑暗处理;B:高温处理;C:NaCl处理。红色、蓝色和黄色分别代表上调、下调和没有表达差异的基因。Figure 5. Expression analyses of LpVQs under dark, high temperature and NaCl treatmentsA: Dark treatment; B: Heat treatment; C: NaCl treatment. Red, blue, and yellow represent up-regulated, down-regulated, and no differential genes, respectively.42 ℃高温处理后,LpVQ的表达模式被分为3类。LpVQ18、LpVQ8、LpVQ28、LpVQ2、LpVQ7、LpVQ4及LpVQ12受1 h高温处理的显著诱导,而高温处理24 h后,其表达又显著下调。LpVQ9、LpVQ22、LpVQ15、LpVQ3、LpVQ42、LpVQ6、LpVQ10、LpVQ1、LpVQ38的表达量在1 h和24 h高温处理后均显著下降。LpVQ5、LpVQ27、LpVQ34、LpVQ14和LpVQ44的表达量只在24 h高温处理后下降(图5:B)。
NaCl处理3 h后,13个LpVQ基因显著上调表达,其中LpVQ4的表达上升了13倍。而NaCl处理38 h后,大部分基因的表达明显下调。仅有3个基因的表达受盐诱导持续上调,分别是 LpVQ8、LpVQ10和LpVQ38(图5:C)。
3. 讨论
近年来,VQ基因已经在多种植物中被发掘鉴定,包括拟南芥、水稻(Oryza sativa L.)、大豆和葡萄(Vitis vinifera L.)等[16-19],而关于多年生黑麦草VQ家族基因的鉴定和分析还鲜见报道。本研究通过全基因组鉴定,得到52个多年生黑麦草LpVQ基因,并系统分析了其理化特性、蛋白结构、基因结构、共线性、进化关系以及响应逆境胁迫的表达模式。
基因结构分析发现,88.46%的LpVQ基因无内含子,与之类似,在拟南芥[16]、水稻[17]、玉米(Zea mays L.)[20]和马铃薯(Solanum tuberosum L.)[21]中,无内含子的VQ基因分别占88.2%、92.5%、88.5%和91.89%。而苔藓(Bryophyta)[22]中无内含子的VQ基因比例则远低于以上物种,仅为28.0%,说明VQ基因可能在长期进化过程中丢失了内含子。
系统进化分析结果显示,多年生黑麦草和拟南芥的VQ家族成员可被分为7个亚家族,且LpVQs与AtVQs亲缘关系较远,只存在4对直系同源基因,分别是LpVQ6 / AtVQ7、LpVQ11 / AtVQ9、LpVQ50 / AtVQ32和LpVQ5 / AtVQ31。根据拟南芥VQ基因的功能,可以推测多年生黑麦草中其直系同源VQ基因的功能。有研究指出,AtVQ9作为AtWRKY8的抑制子,能提高拟南芥的耐盐性[9],由此推测,其直系同源基因LpVQ11可能也在多年生黑麦草抵御盐胁迫中发挥重要作用。
VQ基因在植物响应非生物胁迫中发挥着重要作用。如,过表达PtVQ1基因显著增强了转基因拟南芥的耐盐性[23];而过表达MdVQ37则提高了转基因苹果(Malus pumila Mill.)对高温胁迫的敏感性[24]。本研究对黑暗、高温及盐胁迫处理下多年生黑麦草的VQ表达量进行了分析。结果显示,LpVQ基因广泛响应黑暗、高温及盐胁迫,且大部分LpVQ基因均在胁迫初期即做出快速响应,这与西葫芦(Cucurbita pepo L.)[8]、黄瓜(Cucumis sativus L.)[25]等物种中的研究结果相类似。然而,LpVQ基因的表达模式在不同胁迫处理下也存在差异。如,LpVQ1在黑暗胁迫处理后持续下调表达,而在高温和盐胁迫下持续上调表达,推测该基因可能在不同胁迫中发挥着不同的作用。尽管LpVQ可能参与多年生黑麦草对黑暗、高温及盐等胁迫响应的调控,但其具体功能还需要进一步验证。
综上所述,本研究对多年生黑麦草VQ基因家族成员进行了系统鉴定与分析,为后续验证VQ家族成员在多年生黑麦草应答生物或非生物胁迫中的功能提供了基础。
1 1) 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
图 1 洪平杏及其近缘种叶及果核形态特征
A、A’、E:洪平杏(Armeniaca hongpingensis);B、B’、F:杏(A. armeniaca);C、C’、G:梅(A. mume);D、D’、H:杏梅(A. mume var. bungo)。
Figure 1. Morphological characters of leaves and kernels
![]()
图 2 洪平杏及近缘种的PAM和UPGMA聚类分析
Figure 2. PAM and UPGMA cluster analysis of Armeniaca hongpingensis and related species
![]()
图 3 叶背表皮毛 (A~D)、气孔(E~H) 、叶正面表皮细胞(I~L)及花粉 (M~T)电镜扫描图
A、E、I、M、Q:洪平杏(Armeniaca hongpingensis);B、F、J、N、R:杏(A. vulgaris);C、G、K、O、S:梅(A. mume);D、H、L、P、T:杏梅(A. mume var. bungo)。
Figure 3. Morphological characters of trichome (A–D), stomata (E–H) on leaf abaxial surface, epidermal cell on leaf adaxial surface (I–L), and pollen (M–T)
表 1 洪平杏及其近缘种定性性状及其赋值
Table 1 Selected characters and values for Armeniaca hongpingensis and related species
性状
Characters性状分类及赋值
Classification and values of characters一年生枝颜色 浅绿色—0 红褐色—1 紫红色—2 深紫色—3 一年生枝被毛 无—0 有—1 叶形 宽卵圆形—0 椭圆到椭圆状卵形—1 卵圆形—2 叶尖 短渐尖至急尖—0 长渐尖至尾尖—1 尾尖—2 叶基 宽楔形至圆形—0 圆形至近心形—1 叶缘 细锐锯齿—0 圆钝锯齿—1 叶柄腺体 无—0 有—1 叶片正面被毛 无—0 有—1 叶片背面被毛 无—0 有(密被浅黄褐色柔毛)—1 叶脉被毛 无或具短柔毛—0 密被浅黄褐色柔毛—1 果实被毛 微被短柔毛—0 密被浅黄褐色柔毛—1 核形 卵圆形—0 圆形—1 核顶端 圆钝—0 小钝尖—1 急尖—2 核表面 光滑或稍粗糙—0 蜂窝状孔穴—1 核腹面 龙骨状棱—0 纵沟—1  下载: 导出CSV
下载: 导出CSV
表 2 洪平杏、梅、杏梅及杏的数量性状
Table 2 Morphological quantitative characters of Armeniaca hongpingensis, A. mume, A. mume var. bungo, and A. vulgaris (mean±SD)
性状
Characters / mm洪平杏
A. hongpingensis杏
A. vulgaris梅
A. mume杏梅
A. mume var. bungo叶长 121.22±21.04a 64.49±11.52d 79.22±8.71c 108.86±6.22b 叶宽 57.34±11.37a 50.12±8.49b 41.52±4.63c 61.12±4.94a 叶柄长 17.15±1.74c 24.73±4.92a 11.14±1.58d 18.67±2.18b 叶长/宽 2.13±0.15a 1.29±0.10d 1.91±0.16b 1.79±0.12c 叶柄长/叶长 0.15±0.03c 0.39±0.06a 0.14±0.02d 0.17±0.02b 花冠直径 35.61±2.48a 25.95±1.20c 24.14±0.87d 31.46±2.21b 花梗长 4.19±0.72a 1.33±0.25d 3.47±0.39b 2.70±0.23c 花瓣长 19.09±1.18a 13.00±0.70c 12.14±1.06d 13.89±1.00b 花瓣宽 14.99±1.57a 12.23±0.94b 10.50±1.10c 14.58±1.11a 萼片长 5.11±0.40a 5.05±0.43a 4.43±0.32b 5.07±0.32a 萼片宽 3.55±0.31c 4.53±0.42a 3.73±0.29bc 3.84±0.33b 果长 33.52±1.71c 34.18±3.32a 33.9±1.64b 29.89±1.64c 果宽 33.52±1.70b 35.28±1.96a 28.1±2.13c 35.88±3.76a 果厚 30.55±1.53b 35.2±2.04a 26.5±1.65c 32.10±4.13b 果长宽比 1.09±0.04a 0.096±0.03c 1.04±0.05b 0.99±0.06c 果宽厚比 1.10±0.04a 1.00±0.04c 1.06±0.04b 1.12±0.06a 核长 25.12±1.66a 20.66±0.83c 20.85±2.16c 23.36±1.88b 核宽 17.36±1.19b 16.25±0.58c 14.22±1.32d 19.6±1.55a 核厚 10.27±0.68c 9.52±0.4d 11.23±0.73b 15.09±1.15a 核长宽比 1.45±0.10a 1.27±0.03b 1.48±0.18a 1.19±0.05c 核宽厚比 1.69±0.09a 1.71±0.05a 1.26±0.07b 1.30±0.04b 注:同行不同字母代表差异显著(P<0.05)。下同。 Note: Different letters on same line represent significant differences (P<0.05). Same below.
下载: 导出CSV
表 3 杏属4种植物的叶表面气孔及条纹数量特征
Table 3 Stomata and stripe quantitative characters on leaf abaxial surface of four Armeniaca species
性状Characters 洪平杏
A. hongpingensis杏
A. vulgaris梅
A. mume杏梅
A. mume var. bungo气孔长 / μm 20.96±2.02a 28.00±2.47b 20.92±2.03a 20.67±1.11a 气孔宽 / μm 15.82±1.04a 22.15±2.21c 17.06±1.41b 17.10±1.32b 气孔密度 / ind/mm2 549.25±114.02a 361.00±93.45c 809.63±159.25b 809.25±88.73b 条纹数 11.18±2.19a 8.61±2.40c 18.39±5.30b 9.32±2.30c 条纹宽 / μm 0.84±0.13a 1.29±0.16c 0.67±0.13b 0.87±0.12a
下载: 导出CSV
表 4 杏属4种植物的花粉形态、大小及表面条纹比较
Table 4 Comparison of pollen morphology, size, and stripes among four species of Armeniaca
性状
Characters洪平杏
A. hongpingensis杏
A. vulgaris梅
A. mume杏梅
A. mume var. bungo花粉形态 长球形 长球形 长球形 长球形 对称性与极性 等级辐射对称 等级辐射对称 等级辐射对称 等级辐射对称 外壁纹饰 间断粗条纹,走向不规则 细条纹,走向规则 粗条纹,走向规则 粗条纹,走向略不规则 极轴长(P)/ μm 33.08±1.62a 33.61±1.75a 31.18±1.13b 32.54±1.47ab 赤道轴长(E) / μm 24.59±0.76a 23.21±1.50c 20.50±1.56b 23.00±1.34c P/E 1.35±0.09a 1.45±0.13bc 1.53±0.12b 1.42±0.12ac 条纹宽 / μm 0.34±0.07a 0.16±0.03b 0.35±0.04a 0.34±0.04a
下载: 导出CSV
-
[1] Whitney KD,Ahern JR,Campbell LG,Albert LP,King MS. Patterns of hybridization in plants[J]. Perspect Plant Ecol Evol Syst,2010,12(3):175−182. doi: 10.1016/j.ppees.2010.02.002
[2] Lu LD, Bruce B. Armeniaca Scopoli[M]//Wu ZY, Raven PH, Hong DY, eds. Flora of China: Vol. 9. Beijing: Science Press, 2003: 396-401.
[3] Mehlenbacher SA,Cociu V,Hough FL. Apricots (Prunus)[J]. Genet Resour Temper Fruit Nut Crops,1991,290:65−110.
[4] Byrne DH. Isozyme phenotypes support the interspecific hybrid origin of Prunus xdasycarpa Ehrh[J]. Fruit Var J,1993,47(3):143−145.
[5] Li M,Zhao Z,Miao XJ. Genetic diversity and relationships of apricot cultivars in North China revealed by ISSR and SRAP markers[J]. Sci Hortic,2014,173:20−28. doi: 10.1016/j.scienta.2014.04.030
[6] Shimada T,Haji T,Yamaguchi M,Takeda T,Nomura K,Yoshida M. Classification of Mume (Prunus mume Sieb. et Zucc. ) by RAPD assay[J]. J Japan Soc Hort Sci,1994,63(3):543−551. doi: 10.2503/jjshs.63.543
[7] 中国科学院植物志编辑委员会. 中国植物志: 第38卷[M]. 北京: 科学出版社, 1986: 24-33. [8] 陈俊愉,包满珠. 中国梅(Prunus mume)的植物学分类与园艺学分类[J]. 浙江林学院学报,1992,9(2):119−132. Chen JY,Bao MZ. Botanical classification and Horticultural classification of Chinese Mei (Prunus mume) resources[J]. Journal of Zhejiang Forestry College,1992,9(2):119−132.
[9] 包满珠. 我国川、滇、藏部分地区野梅种质资源及梅的系统学研究[D]. 北京: 北京林业大学, 1991: 1-10. [10] 王家琼. 中国杏属植物花粉形态研究、分布及其亲缘关系研究[D]. 广州: 华南农业大学, 2010: 17-28. [11] 王家琼,吴保欢,崔大方,羊海军,黄峥,齐安民. 基于30个形态性状的中国杏属(Armeniaca Scop. )植物分类学研究[J]. 植物资源与环境学报,2016,25(3):103−111. doi: 10.3969/j.issn.1674-7895.2016.03.13 Wang JQ,Wu BH,Cui DF,Yang HJ,Huang Z,Qi AM. Taxonomic study on Armeniaca Scop. species in China based on thirty morphological characters[J]. Journal of Plant Resources and Environment,2016,25(3):103−111. doi: 10.3969/j.issn.1674-7895.2016.03.13
[12] 刘志娥,王春晖,刘玮琦,汪小凡. 基于核基因和叶绿体基因序列的杏属系统发育分析——探讨洪平杏的起源和亲缘关系[J]. 植物科学学报,2018,36(5):633−641. Liu ZE,Wang CH,Liu WQ,Wang XF. Molecular phylogeny of Armeniaca based on nuclear and chloroplast gene sequences:Exploring the origin and genetic relationship of Armeniaca hongpingensis[J]. Plant Science Journal,2018,36(5):633−641.
[13] Walker JW. Evolution of exine structure in the pollen of primitive angiosperms[J]. Am J Bot,1974,61(8):891−902. doi: 10.1002/j.1537-2197.1974.tb12315.x
[14] Salmaki Y,Jamzad Z,Zarre S,Bräuchler C. Pollen morphology of Stachys (Lamiaceae) in Iran and its systematic implication[J]. Flora Morphol Distrib Funct Ecol Plants,2008,203(8):627−639.
[15] Song JH,Moon HK,Hong SP. Pollen morphology of the tribe Sorbarieae (Rosaceae)[J]. Plant Syst Evol,2016,302(7):853−869. doi: 10.1007/s00606-016-1303-9
[16] Lechowicz K,Wrońska-Pilarek D,Bocianowski J,Maliński T. Pollen morphology of Polish species from the genus Rubus L. (Rosaceae) and its systematic importance[J]. PLoS One,2020,15(5):e0221607. doi: 10.1371/journal.pone.0221607
[17] Xiong XH,Zhou XM,Li M,Xu B,Deng HN,et al. Pollen morphology in Rubus (Rosaceae) and its taxonomic implications[J]. Plant Syst Evol,2019,305(8):705−716. doi: 10.1007/s00606-019-01600-7
[18] 廖明康,张平,郭丽霞,车凤斌. 新疆杏属植物花粉形态的观察[J]. 西北农 业学报,1994,3(4):13−16. Liao MK,Zhang P,Guo LX,Che FB. An observation of Xinjiang apricot pollen morphology[J]. Acta Agriculturae Boreali-Occidentalis Sinica,1994,3(4):13−16.
[19] 杨会侠. 中国杏属植物花粉形态研究[D]. 沈阳: 沈阳农业大学, 2000: 6-21. [20] Arzani K,Nejatian MA,Karimzadeh G. Apricot (Prunus armeniaca) pollen morphological characterisation through scanning electron microscopy,using multivariate analysis[J]. N Z J Crop Hortic Sci,2005,33(4):381−388. doi: 10.1080/01140671.2005.9514374
[21] Erdtman G. Handbook of Palynology: Morphology, Taxonomy, Ecology[M]. New York: Hafner, 1969: 1-486.
[22] Erdtman G. Pollen Morphology and Plant Taxonomy: Angiosperms: Vol. 1[M]. Stockholm: Almqvist och Wiksell, 1952: 1-539
[23] Yilmaz KU, Gurcan K. Genetic diversity in apricot[M]//Caliskan M, ed. Genetic Diversity in Plants. Rijeka: In Tech, 2012: 297-301.
[24] Hamzeh’ee B,Attar F,Assareh MH,Maassoumi AA,Kazempour S. Comparative micromorphological characteristics of lower surface of leaf epidermis and seed surface in two series of Crataegus l. (Rosaceae) and their taxonomical role[J]. Iran J Bot,2016,22(2):125−137.
[25] Attar F,Esfandani-Bozchaloyi S,Mirtadzadini M,Ullah F,Zaman W. Foliar and stem epidermal anatomy of the tribe Cynoglosseae(Boraginaceae) and their taxonomic significance[J]. Microsc Res Tech,2019,82(6):786−802. doi: 10.1002/jemt.23223
[26] Ullah F,Ayaz A,Saqib S,Parmar G,Bahadur S,Zaman W. Taxonomic implication of leaf epidermal anatomy of selected taxa of Scrophulariaceae from Pakistan[J]. Microsc Res Tech,2021,84(3):521−530. doi: 10.1002/jemt.23608
[27] Yang HQ,Wang H,Li DZ. Comparative morphology of the foliage leaf epidermis,with emphasis on papillae characters,in key taxa of woody bamboos of the Asian tropics (Poaceae:Bambusoideae)[J]. Bot J Linn Soc,2008,156(3):411−423. doi: 10.1111/j.1095-8339.2007.00736.x
[28] Zhang YX,Zeng CX,Li DZ. Scanning electron microscopy of the leaf epidermis in Arundinarieae (Poaceae:Bambusoideae):evolutionary implications of selected micromorphological features[J]. Bot J Linn Soc,2014,176(1):46−65. doi: 10.1111/boj.12192
[29] Geng Z,Liu B,Rohwer JG,Ferguson DK,Yang Y. Leaf epidermal micromorphology defining the clades in Cinnamomum (Lauraceae)[J]. PhytoKeys,2021,182:125−148. doi: 10.3897/phytokeys.182.67289
[30] Song JH,Hong SP. Taxonomic significance of the leaf micromorphology in the tribe Sorbarieae (Spiraeoideae:Rosaceae)[J]. Korean J Plant Taxon,2016,46(2):199−212. doi: 10.11110/kjpt.2016.46.2.199
[31] Babosha A,Ryabchenko A,Kumachova T,Komarova G,Yatsenko I. Micromorphology of the leaf surface in some species of Dryadoideae (Rosaceae)[J]. Micron,2023,167:103428. doi: 10.1016/j.micron.2023.103428
[32] Khadgi A,Weber CA. Morphological characterization of prickled and prickle-free Rubus using scanning electron microscopy[J]. HortScience,2020,55(5):676−683. doi: 10.21273/HORTSCI14815-20
[33] 刘有春,陈伟之,刘威生,刘宁,张玉萍,刘硕. 仁用杏起源演化的孢粉学研究[J]. 园艺学报,2010,37(9):1377−1387. Liu YC,Chen WZ,Liu WS,Liu N,Zhang YP,Liu S. Palynological study on the origin and systematic evolution of kernel-using apricots[J]. Acta Horticulturae Sinica,2010,37(9):1377−1387.
[34] 韩雪平,薛晓敏,王金政,王贵平. 基于花粉微观特征的5个杏品种孢粉学研究[J]. 江苏农业科学,2018,46(14):124−127. Han XP,Xue XM,Wang JZ,Wang GP. Study on palynology of five apricot cultivars based on pollen microscopic characteristics[J]. Jiangsu Agricultural Sciences,2018,46(14):124−127.
[35] 董英山,郝瑞. 西伯利亚杏、普通杏及东北杏亲缘关系探讨[J]. 吉林农业大学学报,1991,13(1):24−27. Dong YS,Hao R. Studies of the sibship of Prunus sibirica L.,Prunus armeniaca L. and Prunus mandshrica Koehne[J]. Journal of Jilin Agricultural University,1991,13(1):24−27.
-
其他相关附件
-
PDF格式
刘志娥附表1 点击下载(162KB)
-






计量
- 文章访问数: 303
- HTML全文浏览量: 34
- PDF下载量: 148
