
Effects of intercropping forage on weed community composition and diversity in an apple orchard in the river valley area of Nyingchi, Tibet
-
摘要:
以林芝市巴宜区苹果园杂草群落为研究对象,以清耕后自然生长植物为对照,研究了一年生箭筈豌豆(Avena sativa L.)、燕麦(Vicia sativa L.)和多年生紫花苜蓿(Medicago sativa L.)、鸭茅(Dactylis glomerata L.)牧草单播或混播对苹果园杂草群落组成和多样性的影响。结果显示:(1)调查期间共发现杂草17科31属33种,其中优势科为禾本科、豆科、蓼科和菊科,优势种为白草(Pennisetum flaccidum Griseb.)、画眉草(Eragrostis pilosa (L.) Beauv.)、小头蓼(Persicaria microcephala (D. Don) H. Gross.)、牛膝菊(Galinsoga parviflora Cav.)和蒲公英(Taraxacum mongolicum Hand.-Mazz.);(2)牧草种植可以减少果园中杂草种类,降低优势杂草重要值,改变杂草群落结构,不同类型牧草在不同种植年限对杂草群落的影响存在差异;(3)不同间作牧草模式下杂草群落物种多样性有显著差异,但整体上较对照有所降低,种植年限与种植模式对群落多样性和优势度均有显著影响;(4)间作牧草有良好的杂草抑制效果,其中禾本科牧草的杂草防控效果优于豆科牧草,混播处理的杂草防控效果优于单播处理。研究结果表明,人工牧草的选择应考虑牧草生长年限,一年生牧草在短期内杂草防控效果较好,多年生牧草适宜长期种草的果园。
Abstract:To study the effects of intercropping with forage grasses on the composition and diversity of weed communities in an apple orchard in Nyingchi, Tibet, two annual grasses (Avena sativa L. and Vicia sativa L.) and two perennial grasses (Medicago sativa L. and Dactylis glomerata L.) were single sown or mixed sown, with grasses naturally growing after clear tillage used as a control. The weed communities were then surveyed in 2011 and 2022, respectively. A total of 33 weed species belonging to 31 genera and 17 families were found in the experimental plots in the two studied years. The dominant families were Gramineae, Leguminosae, Polygonaceae, and Asteraceae, and the dominant species were Pennisetum flaccidum Griseb., Eragrostis pilosa (L.) Beauv., Galinsoga parviflora Cav., Taraxacum mongolicum Hand.-Mazz., and Persicaria microcephala (D. Don) H. Gross. Intercropping with forage grasses led to a reduction in weed species richness and importance values of the dominant weed species, which were influenced by forage grass species and planting year. Intercropping also resulted in lower species diversity of the weed communities, with both planting year and cropping pattern significantly affecting community diversity and dominance. Weed control efficacy was higher for Gramineae forage compared to Leguminosae forage, and mixed sowing showed better results than single sowing treatment. The above findings suggest that the selection of forage grasses should be based on their growth period, with annual forages being more effective for short-term weed control and perennial forages being more suitable for long-term grass planting in orchards.
-
Keywords:
- Tibet /
- Intercropping /
- Forage grass /
- Species diversity /
- Weed control /
- Weed community
-
西藏位于我国五大传统牧区之一的青藏高原,平均海拔达到4 000 m,内陆冰川和高山湖泊众多,高海拔和恶劣的自然条件使得西藏北部形成了独特的高寒草地生态系统[1]。近年来随着人类活动的增加与气候变化的加剧,西藏地区天然草地大量退化,同时,随着畜牧业的不断发展,西藏畜牧业逐步由放牧转型为畜舍饲养,西藏地区对于人工牧草的需求也越来越高[2]。2016年青藏高原约有绵羊84万只,牦牛505万头,但在2015年,西藏的“一江两河”流域饲草短缺就有近50.4万吨[3],草畜矛盾亟待解决[4]。
林芝地处西藏一江两河流域,自然资源丰富,环境适宜,是西藏南草北调政策中的重要牧草产区。随着近年来交通的便利以及政府对水果产业的扶持,西藏一江两河流域的果园产业发展迅速:1990年西藏全区苹果园面积大约为508 hm2,产量约为3 696 t,2019年面积达到了2 675 hm2,产量达9 951 t [5]。充分利用果园林下土地种植人工牧草,一方面能充分利用林下边际土地,缓解草畜矛盾,另一方面也会带来额外的经济和生态效益。
果园间种牧草,在牧草盛花期刈割牧草覆盖地表,可以有效地改善土壤理化性质,如增加土壤含水率,提高土壤有机质含量[6],提高土壤速效养分如有效磷、碱解氮、速效钾的含量[7],调节土壤pH值,从而改良盐碱地土壤等[8];果园生草也会提高果实产量,改善果实品质,改善果园局部小气候[9]。日常农业生产过程中,往往伴随杂草生长,尤其是恶性杂草的大量生长,会与果树和牧草争水争肥,甚至竞争生长空间,从而降低果树和牧草的产量和品质。杂草相较于农作物,有着更强的生长速度和养分利用能力,无人工干预时,杂草通常要比农作物生长更加茂盛[10]。因此,果园杂草防治是果园管理的重要环节之一。人工合理种植牧草可以通过占领生态位来控制杂草生长[11]。肖润林等[12]的研究表明,在茶园间作白三叶草(Trifolium repens L.),能够降低优势集中性指数,有效控制了园内优势恶性杂草的生长。王加国等[13]研究发现,猕猴桃园种植人工牧草后,牧草会占据地表大部分空间,从而抑制杂草的萌发与生长。王晶晶等[14]的研究表明,在蓝莓园间作白三叶草,对杂草的防控效果达80%以上,极大减少杂草防治成本。目前关于“以草治草”的研究较多,但有关林芝地区果园杂草种类,以及牧草不同间作模式对杂草的控制效果的研究还较少,缺乏系统研究。
本研究以林芝河谷区苹果园为试验区,在园内人工种植箭筈豌豆(Avena sativa L.)、燕麦(Vicia sativa L.)、紫花苜蓿(Medicago sativa L.)以及鸭茅(Dactylis glomerata L.),研究几种牧草的不同单、混播模式对苹果园内杂草群落特征的影响,以期为西藏河谷区人工牧草种植模式筛选以及果园杂草管理提供科学依据和参考。
1. 材料与方法
1.1 试验地概况
试验地位于西藏自治区林芝市巴宜区布久乡杰麦村藏汉情缘苹果园(29°34'14.73" N,94°25'56.10" E),海拔3 005 m,属高原温带湿润季风气候,气候温和,平均日照数2 022 h以上,年均气温8.7 ℃,无霜期长达180 d,生长期集中于5-10月。年降水量约650 mm,降水集中于夏季(6-8月)。试验地栽种苹果(Malus pumila Mill.)品种为‘黑钻苹果’,树龄15年,树高3 m,冠幅2.5 m × 2 m,行距4 m,株距3 m,处于盛果期。河谷地貌,土壤质地为粉砂壤土,耕作层厚度20 cm,全氮全碳较低,速效磷钾较丰富。
1.2 试验设计
本试验设计包含一年生豆科(箭筈豌豆)、禾本科(燕麦)牧草单播和混播,多年生豆科(紫花苜蓿)、禾本科(鸭茅)牧草单播和混播,共计6种模式,并以清耕后自然生长植物作为对照,共7种处理(表1),每个处理3次重复。试验采用随机区组设计,共21个小区,每个小区包括果树6行,每行5株,面积为300 m2,树盘以外行间种草。牧草播种时间为2021年5月中旬,其中一年生牧草每年5月重新播种,多年生牧草仅在试验第一年播种,混播播种量为单播播种量的70%(表1)。每年9月盛花期对所有处理割草一次,将割下的草均匀覆盖于果园行间。试验地块的地形、生态环境一致,日常管理按照果园原有养护方案进行。
表 1 不同处理牧草播种量Table 1. Sowing of forages in different treatments处理
Treatment简称
Abbreviation牧草品种
Forage species播种量
Seeding volume / g/m2一年生豆科单播 L1 箭筈豌豆Avena sativa L. 15 一年生禾本科单播 G1 燕麦Vicia sativa L. 30 一年生豆禾混播 L1G1 箭筈豌豆、燕麦 10.5、21 多年生豆科单播 L2 紫花苜蓿Medicago sativa L. 1.5 多年生禾本科单播 G2 鸭茅Dactylis glomerata L. 3 多年生豆禾混播 L2G2 紫花苜蓿、鸭茅 1.05、2.1 对照 CK − − 1.3 群落调查
分别于2021年和2022年8月中旬进行杂草群落调查,采用随机样方法,每个小区随机设置3个1 m × 1 m的样方,记录样方内所有杂草的种类、高度、盖度及频度。
1.4 数据处理
采用重要值分析群落内杂草的优势度,植物群落物种多样性特征用丰富度指数、优势度指数、多样性指数和均匀度指数表示,分别为Patrick指数、Simpson指数、Shannon-Wiener指数以及Pielou指数。各指标的计算方式如下:
(1)重要值(Fi 、Ai、Ci分别为第i个物种的相对频度、相对多度和相对盖度):
P=(Fi+Ai+Ci)/3×100% (2)Patrick指数:R = S(S为样方中所有物种数目)
(3)Simpson指数(Ni为第i个物种的数量,N为所有物种总个体数):
λ=S∑i=1Ni(Ni−1)N(N−1) (4)Shannon-Wiener指数(Pi为第i个物种的重要值):
H′e=−S∑i=1PilnPi (5)Pielou指数:
Je=H′H′max=H′lnS 杂草的抑制效果根据杂草的总相对盖度来确定,总相对盖度为各处理中所有杂草相对盖度的累加值,杂草总相对盖度越小代表杂草防除效果越好。
不同处理间群落的物种多样性指数和总相对盖度的差异采用单因素方差分析(One-way ANOVA);种植模式、种植年限对物种多样性指数的影响使用多因素方差分析(Two-way ANOVA);以各处理中重要值大于5%的杂草物种与处理构成矩阵进行系统聚类分析,确定不同间作牧草处理后杂草群落结构的相似程度。采用SPSS 26.0软件进行数据分析。
2. 结果与分析
2.1 不同牧草间作模式对苹果园杂草组成及重要值的影响
根据样方调查数据统计结果(表2、表3),2021年和2022年本研究在试验地内共发现杂草17科31属33种,主要为禾本科、豆科、蓼科和菊科杂草。其中2021年试验地内有杂草14科25属26种,2022年有杂草14科24属24种。从生长周期来看,2021年调查中的杂草以一年生杂草为主,共计17种,占比65.4%。2022年调查杂草中一年生杂草共14种,占比58.3%。从所调查的物种组成来看,2021年白草(Eragrostis pilosa (L.) Beauv.)、马唐(Digitaria sanguinalis (L.) Scop.)、牛膝菊(Galinsoga parviflora Cav.)、蒲公英(Taraxacum mongolicum Hand.-Mazz)、小头蓼(Persicaria microcephala (D. Don) H. Gross)、野苜蓿(Medicago falcata L.)、灰绿藜(Oxybasis glauca (L.) S. Fuentes, Uotila & Borsch)、菊叶香藜(Dysphania schraderiana (Roemer & Schultes) Mosyakin & Clemants)和野葵(Malva verticillata L.)在多个处理中均有出现,说明其在本试验果园中普遍存在。广布野豌豆(Vicia cracca L.)、白车轴草(Trifolium repens L.)、拉拉藤(Galium spurium L.)、黄苞南星(Arisaema flavum (Forsk.) Schott)等仅出现在1个样方中,重要值均较低,为偶见种。2022年,画眉草(Eragrostis pilosa (L.) Beauv.)、白草、牛膝菊、蒲公英、小头蓼、牻牛儿苗(Erodium stephanianum Willd.)和一年蓬(Erigeron annuus (L.) Pers.)等在多个处理中出现,为常见种,在果园普遍存在。鹅肠菜(Stellaria aquatica (L.) Scop.)、毛车前(Plantago australis Lam.)、灯芯草(Juncus effusus L.)和广布野豌豆仅在少量样方中出现,为偶见种。从两年调查的结果来看,2022年杂草科属数与2021年基本无差异,杂草种类少于2021年,一年生杂草种类所占比例有所下降,偶见种数量减少,出现了宝盖草(Lamium amplexicaule L.)、紫花香薷(Elsholtzia argyi Lévl.)、毛车前、鹅肠菜等新的杂草种类。
表 2 2021年不同处理中杂草群落组成及重要值Table 2. Weed community composition and importance values in different treatments in 2021科名
Family种名
Species处理 Treatment CK L1 G1 L1G1 L2 G2 L2G2 禾本科 画眉草 Eragrostis pilosa (L.) Beauv. 14.55 4.32 3.14 − 20.83 3.90 2.63 白草 Pennisetum flaccidum Griseb. 32.87 17.26 6.02 3.76 14.57 21.19 26.15 马唐 Digitaria sanguinalis (L.) Scop. − − − − − − 4.90 菊科 牛膝菊 Galinsoga parviflora Cav. 8.44 8.51 3.18 6.60 12.83 8.04 10.28 猪毛蒿 Artemisia scoparia Waldst. et Kit. 0.76 − − 0.80 − 0.77 − 蒲公英 Taraxacum mongolicum Hand.-Mazz. 4.75 3.08 6.27 2.66 6.16 5.12 2.50 秋鼠曲草 Pseudognaphalium hypoleucum(Candolle) Hilliard & B. L. Burtt 0.85 − − − − − 0.70 蓼科 小头蓼 Persicaria microcephala (D. Don) H. Gross 12.79 13.83 7.13 8.70 7.48 6.45 5.21 水蓼 Persicaria hydropiper (L.) Spach 0.95 − − − − 0.51 − 萹蓄 Polygonum aviculare L. 0.62 − 0.63 − 1.55 0.53 − 豆科 广布野豌豆 Vicia cracca L. − − − − − − 0.62 野苜蓿 Medicago falcata L. 5.96 3.24 2.99 0.62 1.41 8.16 0.74 白车轴草 Trifolium repens L. − − − − 4.59 − − 蔷薇科 绢毛委陵菜 Potentilla sericea L. 0.57 − − − 1.04 0.76 0.65 蛇莓 Duchesnea indica (Andr.) Focke − − − 0.69 − − 0.50 苋科 灰绿藜 Oxybasis glauca (L.) S. Fuentes, Uotila & Borsch 2.37 3.26 2.55 − 2.00 3.32 1.99 菊叶香藜 Dysphania schraderiana (Roemer & Schultes) Mosyakin & Clemants 0.76 − 1.37 2.64 4.78 1.43 1.92 莎草科 青绿薹草 Carex breviculmis R. Br. − 1.04 − − − − − 扁穗草 Blysmus compressus (L.) Panz. 0.63 − − − − 0.81 1.86 锦葵科 野葵 Malva verticillata L. 3.21 0.76 1.44 3.03 3.59 4.00 6.47 牻牛儿苗科 牻牛儿苗 Erodium stephanianum Willd. 1.17 − 0.64 − 2.10 − − 大麻科 大麻 Cannabis sativa L. 0.58 1.54 − − 2.48 0.53 0.48 天南星科 黄苞南星 Arisaema flavum (Forsk.) Schott 0.57 − − − − − − 灯芯草科 灯芯草 Juncus effusus L. 1.44 − − − − − 1.71 茜草科 拉拉藤 Galium spurium L. − − − − 0.49 − − 木贼科 节节草 Equisetum ramosissimum Desf. 1.30 − 0.63 0.63 − 0.52 3.48 注:处理简称同表1。下同。 Note: Treatments are the same as shown in Table 1. Same below. 表 3 2022年不同处理中杂草群落组成及重要值Table 3. Weed community composition and importance values in different treatments in 2022科名
Family种名
Species处理 Treatment CK L1 G1 L1G1 L2 G2 L2G2 禾本科 画眉草 Eragrostis pilosa (L.) Beauv. 15.06 10.08 3.46 11.89 10.21 1.83 3.19 白草 Pennisetum flaccidum Griseb. 34.58 54.87 15.46 27.84 6.69 − 4.88 菊科 牛膝菊 Galinsoga parviflora Cav. 7.43 6.49 10.94 21.08 1.33 1.72 1.50 蒲公英 Taraxacum mongolicum Hand.-Mazz. 10.01 10.87 15.24 5.19 13.45 7.26 3.04 秋鼠曲草 Pseudognaphalium hypoleucum (Candolle) Hilliard & B. L. Burtt 2.43 1.13 0.94 0.88 1.23 − − 一年蓬 Erigeron annuus (L.) Pers. 4.37 2.23 3.85 3.07 2.61 − − 白莲蒿 Artemisia stechmanniana Bess. 0.77 − − − 2.51 1.78 − 蓼科 尼泊尔酸模 Rumex nepalensis Spreng. 1.55 1.14 0.88 2.70 2.15 − − 小头蓼 Persicaria microcephala (D. Don) H. Gross 0.82 1.14 4.63 2.25 1.36 − − 萹蓄 Polygonum aviculare L. − 1.17 − − − − − 豆科 广布野豌豆 Vicia cracca L. 0.82 − 0.88 − − − − 野苜蓿 Medicago falcata L. 1.52 − − − − − − 蔷薇科 绢毛委陵菜 Potentilla sericea L. − − − 0.94 − 1.73 − 蛇莓 Duchesnea indica (Andr.) Focke − − 0.88 − − − − 唇形科 宝盖草 Lamium amplexicaule L. 2.64 1.13 2.11 1.90 1.51 − − 紫花香薷 Elsholtzia argyi Lévl. 2.37 − − − − − − 锦葵科 野葵 Malva verticillata L. 0.75 − − 0.91 − − 1.54 牻牛儿苗科 牻牛儿苗 Erodium stephanianum Willd. 2.89 2.28 3.76 1.22 − 3.36 2.99 大麻科 大麻 Cannabis sativa L. 2.99 1.18 1.01 − 1.27 − − 灯芯草科 灯芯草 Juncus effusus L. − − − − − 1.71 − 木贼科 节节草 Equisetum ramosissimum Desf. 2.16 − − − − − 1.64 车前科 毛车前 Plantago australis Lam. − − − − 1.26 − − 石竹科 鹅肠菜 Stellaria aquatica (L.) Scop. − 1.14 − − − − − 莎草科 青绿薹草 Carex breviculmis R. Br. 2.47 1.91 − − − − − 2021年CK处理中的优势杂草为白草、画眉草和小头蓼,重要值均在12%以上,它们在牧草间作处理中重要值均有所降低;间作牧草处理中的优势杂草为白草、小头蓼、画眉草与牛膝菊,其中白草是苹果园中生长优势最大的杂草,它在L1、G2与L2G2中重要值较高,最高达32.80%,除G1与L1G1,其他处理中白草的重要值均高于14%。一年生牧草种植对于杂草种类数和重要值影响要大于多年生牧草,其中抑制效果最好的为L1G1处理,群落中杂草种类相较CK减少了50%,画眉草、白草和小头蓼的重要值分别降低了100%、88.6%和32.0%。2022年CK中优势杂草为白草、画眉草和蒲公英,重要值均在10%以上;间作牧草处理中的优势杂草为白草、画眉草、牛膝菊和蒲公英,其中白草在L1、G1、L1G1等处理中重要值在15%以上,最高达到54.87%。种植牧草第2年,一年生牧草对杂草抑制效果较差,多年生牧草对杂草种类和重要值的影响更大,其中G2和L2G2群落中杂草种类数较CK减少了61.1%, G2杂草群落中白草、画眉草和牛膝菊重要值分别降低了100%、87.8%和76.9%,L2G2中3种杂草重要值分别降低了85.9%、78.9%和79.9%。
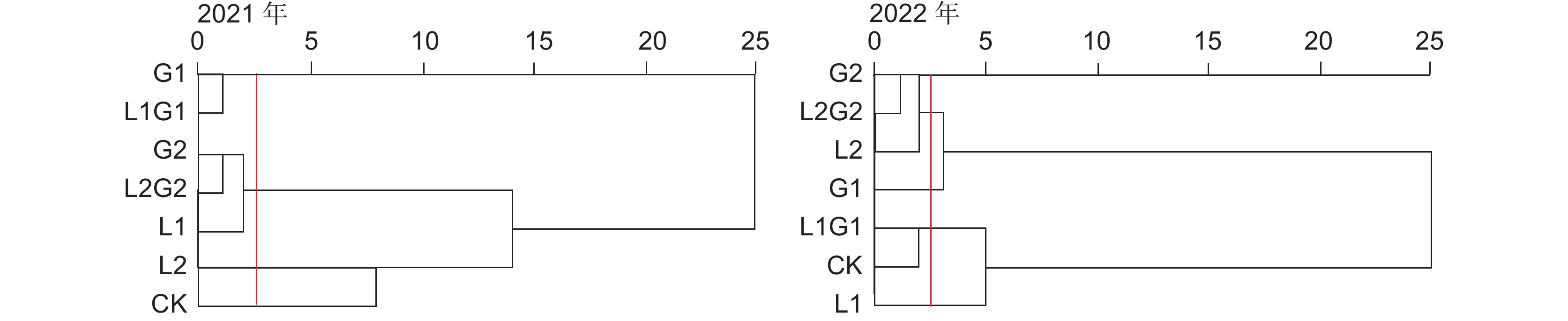
系统聚类分析可以将不同处理按照群落杂草物种及其重要性分类,群落结构相似的处理将归纳为一组。本研究的系统聚类分析结果显示: 2021年7个处理可以划分为4组,G1处理和L1G1处理群落组成类似,G2、L2G2和L1处理群落组成类似,L2、CK处理各为一组;2022年7个处理可以划分为4组,G2、L2G2和L2处理群落组成相似,L1G1和CK处理群落组成相似,G1、L1处理各单独为一组(图1)。
![]() 图 1 不同处理下杂草群落的聚类分析虚线代表在该处对处理进行组别划分。处理简称同表1。下同。Figure 1. Cluster analysis of weed communities in different treatmentsDashed line represents the grouping of treatments at this location. Treatments are the same as shown in Table 1. Same below.
图 1 不同处理下杂草群落的聚类分析虚线代表在该处对处理进行组别划分。处理简称同表1。下同。Figure 1. Cluster analysis of weed communities in different treatmentsDashed line represents the grouping of treatments at this location. Treatments are the same as shown in Table 1. Same below.2.2 间作牧草对苹果园杂草多样性的影响
2.2.1 植物群落丰富度指数和优势度指数分析
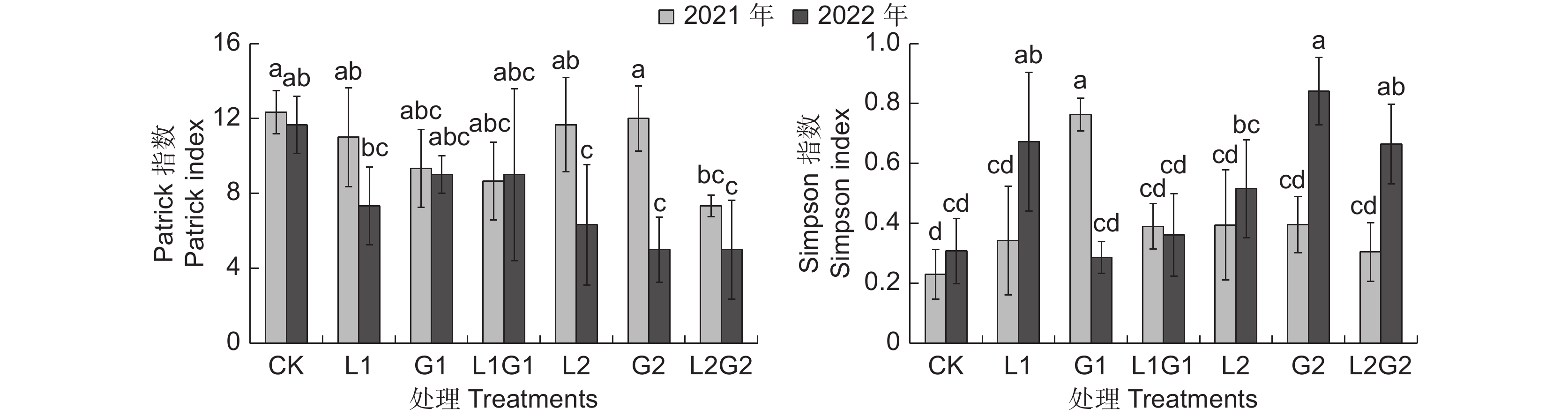
Patrick指数代表群落中物种的种类丰富度,分析结果显示:种草第1年(2021年)苹果园间作牧草处理L1、G1、L1G1、L2、G2和L2G2的Patrick指数分别为11、9、9、12、12和7,其中L2G2的Patrick指数显著低于CK(R=12,P<0.05),且显著低于G2。种草第2年(2022年),6种间作牧草处理的Patrick指数分别为7、9、9、6、5和5,其中L2、G2和L2G2的Patrick指数均显著低于 CK(R=12,P<0.05),间作牧草的处理间则无显著差异(图2)。对比两年数据,CK的Patrick指数变化不显著,但人工种植牧草处理的植物群落的Patrick指数发生了较大改变。相较于2021年,G1 和L1G1的Patrick指数基本不变,而L1、L2、G2和L2G2的Patrick指数分别下降了33.3%、45.7%、58.3%和31.8%,其中L2、G2的Patrick指数降低显著。该结果说明间作牧草第1年,物种丰富度变化不明显,随着种草年限的增加,即间作牧草第2年,多年生牧草处理中的物种丰富度会明显降低。
![]() 图 2 不同处理下植物群落的Patrick指数与Simpson指数不同小写字母表示不同间作牧草处理以及年限间的差异显著(P<0.05)。下同。Figure 2. Patrick and Simpson indices of plant communities in different treatmentsDifferent lowercase letters indicate significant differences at P<0.05 among different treatments in different years. Same below.
图 2 不同处理下植物群落的Patrick指数与Simpson指数不同小写字母表示不同间作牧草处理以及年限间的差异显著(P<0.05)。下同。Figure 2. Patrick and Simpson indices of plant communities in different treatmentsDifferent lowercase letters indicate significant differences at P<0.05 among different treatments in different years. Same below.Simpson指数反映了群落中物种的优势程度,如图2所示,2021年CK、L1、G1、L1G1、L2、G2和L2G2的Simpson指数分别是0.231、0.342、0.763、0.390、0.395、0.395和0.305,其中G1的显著高于CK,而其他间作牧草处理与CK无显著差异;2022年,各处理Simpson指数分别是0.308、0.673、0.287、0.362、0.516、0.841和0.665,L1、G2和L2G2显著高于CK、G1和L1G1。对比两年数据,CK的Simpson指数变化不显著,但种植一年生牧草和多年生牧草后处理的Simpson指数发生了较大改变。相较于2021年,L1、G2和L2G2的Simpson指数增加显著,分别为96.5%、112.9%和118.4%,G1的Simpson指数显著降低了62.4%。该结果说明试验第1年的一年生禾本科单播处理,试验第2年的一年生豆科处理和含有多年生禾本科处理的优势度指数均较高,人工种草显著提高了这些处理中优势种的优势度,且随着种草年限的增加,这些处理的优势种的优势度也在增加。
2.2.2 植物群落多样性和均匀度分析
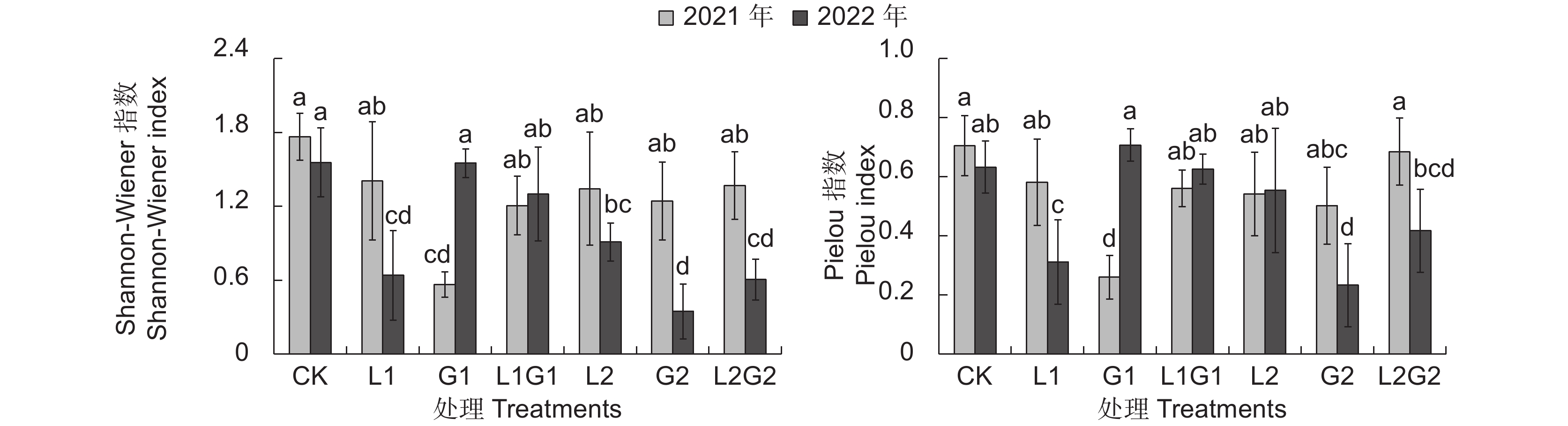
就植物群落的多样性指数而言,种草第1年(2021年)苹果园间作牧草6种处理的Shannon指数分别为1.407、0.565、1.205、1.343、1.243和1.368,其中G1的Shannon指数显著低于CK,且显著低于其他种草处理。种草第2年(2022年),6种间作牧草处理的Shannon指数分别为0.639、1.549、1.301、0.911、0.346和0.604,其中L1、L2、G2和L2G2的Shannon指数均显著低于 CK,G2的Shannon指数显著低于G1、L1G1、L2,L1、L2G2的则显著低于G1、L1G1处理(图3)。相较于2021年,CK和L1G1的Shannon指数变化不明显,其他间作牧草处理的发生了显著变化:L1、L2、G2、L2G2的Shannon指数分别显著下降了54.6%、32.0%、71.8%、56.2%,G1的Shannon指数显著升高了174.1%。该结果说明,人工种草后普遍降低了群落物种的复杂程度,试验第1年种植一年生牧草的影响较为显著,试验第2年种植多年生牧草的影响更明显,杂草多样性显著降低。
![]() 图 3 不同处理下植物群落Shannon指数与Pielou指数Figure 3. Shannon and Pielou indices of plant communities in different treatments
图 3 不同处理下植物群落Shannon指数与Pielou指数Figure 3. Shannon and Pielou indices of plant communities in different treatments均匀度Pielou指数分析结果显示,2021年CK、L1、G1、L1G1、L2、G2和L2G2分别是0.581、0.260、0.560、0.541、0.502和0.685,其中G1的Pielou指数显著低于CK,且显著低于其他种草处理,其他种草处理与CK无显著差异。2022年,各人工种草处理Pielou指数分别是0.312、0.707、0.626、0.554、0.233和0.417,其中L1和G2显著低于CK和其他种草处理。从两年结果对比来看,物种均匀度的变化与多样性的变化极为类似:相较于2021年,CK 和L1G1的Pielou指数变化不明显,间作牧草处理的L1、G2、L2G2的Pielou指数分别显著降低了46.5%、53.5%、39.1%,但G1处理显著增加了171.9%(图3)。该结果说明,试验第1年,人工种草处理的群落中物种分布不均匀,其中单播一年生禾本科处理中均匀度最低;但在试验第2年,单播一年生禾本科处理中物种均匀度增加,而单播多年生禾本科处理和多年生豆禾混播处理的杂草分布较不均匀。
为进一步比较不同处理和种植年份对杂草群落的影响,我们针对种植年份和种植模式对群落多样性进行了双因素方差分析,从表4中可以看出,种植年份对于群落的Patrick指数、Shannon指数、Simpson指数的影响极显著(P<0.01),但对Pielou指数无显著影响;种植模式对Patrick指数的影响显著(P<0.05),对于其他多样性指数作用均为极显著(P<0.01);种植年份和处理类型的交互作用对群落Patrick值无显著影响,对于其他群落指标均有着极显著影响(P<0.01)。
表 4 种植模式和种植年份与植物群落多样性指数的双因素方差检验Table 4. Two-way ANOVA of treatment type and planting year on community diversity index of plant communities影响因子
Impact factor因变量
Dependent
variable自由度
dfF值
F-valueP值
P-value年份 Patrick 1 14.313 0.001 Shannon 1 9.979 0.004 Pielou 1 1.816 0.189 Simpson 1 8.473 0.007 种植模式 Patrick 6 3.211 0.016 Shannon 6 5.254 0.001 Pielou 6 3.997 0.005 Simpson 6 4.335 0.003 年份 × 种植模式 Patrick 6 2.101 0.085 Shannon 6 7.713 0.000 Pielou 6 6.879 0.000 Simpson 6 8.419 0.000 2.3 间作牧草对杂草抑制效果
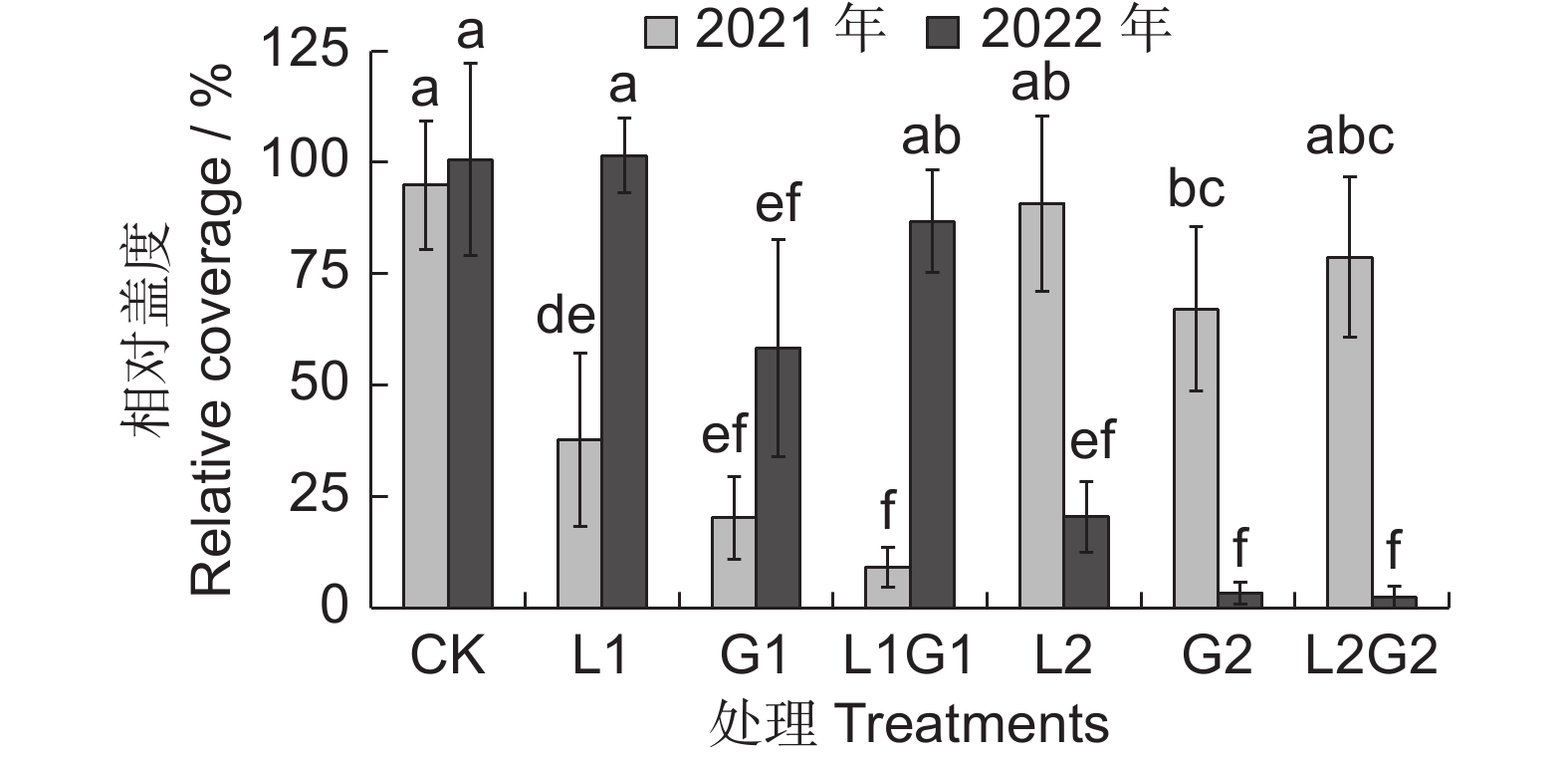
间作牧草处理后杂草的总相对盖度可以反映对杂草的抑制程度。2021年,一年生牧草处理L1、G1、L1G1显著低于CK的杂草群落总相对盖度(94.89%),说明一年生牧草在间作牧草的第一年具有较好的杂草抑制效果。6种间作牧草处理中,杂草相对盖度最高的为L2,达90.74%,最低的L1G1为19.07%,且多年生牧草处理杂草总相对盖度远高于一年生牧草处理。2022年,G1、L2、G2、L2G2等处理杂草群落总相对盖度显著低于CK(100.58%)。6种间作牧草处理中,杂草群落总相对盖度最高的为L1,达101.53%,最低的L2G2为2.36%;一年生牧草处理的杂草总相对盖度均显著高于多年生牧草处理。对比两年结果可看出,CK的杂草相对盖度无明显变化,而间作牧草处理中杂草总相对盖度均出现了大幅度增减:相比于2021年,2022年L1G1的杂草总相对盖度年增加了856.2%,多年生牧草处理群落杂草总相对盖度则大幅降低,其中L2G2减少了97.00%(图4)。
3. 讨论
本研究对西藏自治区林芝市河谷区的苹果园内不同牧草种植模式下的杂草群落进行了调查和分析,2021、2022年共发现杂草17科31属33种,主要为菊科、禾本科、豆科以及蓼科杂草,其中一年生杂草占比分别为65.4%、58.3%。环境条件的变化如人为扰动、土壤含水率等会影响到群落的物种组成[15]。果园中的人为扰动主要有果园土壤的翻耕、水果采摘等,这些活动同时也会对土壤性质产生一定的影响[16]。果园的表层土壤的翻耕会灭除多年生杂草中的浅位性宿根杂草,对于靠种子繁殖的一年生杂草以及部分深位性宿根杂草影响相对较小,且一年生的短命植物更容易在高强度和高频率的人为干扰环境下生长和繁殖[16]。因此,本研究中一年生杂草占比明显高于多年生杂草。本试验区中的优势杂草有白草、画眉草、蒲公英和小头蓼等,其中白草、蒲公英均属于高寒地区农田恶性杂草,它们种子数量多、扩散快,生长迅速,繁殖能力强,对于作物危害较大[17]。白草为深位性宿根杂草,地下根茎发达,多蔓延成网状根茎系统[18],在高原环境下,白草会将更多资源投入地下根部生长,对于环境扰动的抵抗能力进一步加强[19];画眉草、蒲公英和小头蓼为一年生杂草,生长周期短,繁殖快,种子数量多且萌发率高,虽然受人工翻耕影响较大,但庞大的数量使其在苹果园生态系统中仍占据优势地位,因此相对于其他杂草,这几种杂草在苹果园生态系统中存在更广泛。
群落中植物的物种组成是群落植被最直观的特征,能反映群落物种的丰富度和多样性[20]。已有研究表明,人工生草会对群落物种组成产生影响。朱先波等[21]研究发现,在猕猴桃园内人工种植白三叶可以大幅度降低果园内杂草种类,人工种植的牧草会占据一部分生长空间,在生长期与杂草竞争水分和养分,在土地翻耕后直接撒播草种,牧草早发,迅速占据周围资源,后发杂草难以侵入,一方面能减少杂草物种数量,另一方面也会抑制优势杂草的生长。本试验中间作牧草相较于CK显著减少了杂草群落物种数量,抑制了优势杂草生长。第1年土地翻耕后,间作牧草降低了白草、画眉草等杂草的重要值,且种植一年生牧草的处理影响更明显,第2年间作牧草也显著降低了白草、画眉草的重要值,但牛膝菊和蒲公英在群落中的重要值略有提高,这可能与其他杂草牛膝菊和蒲公英一季多代、生长迅速,更能适应环境变化的特性有关[22]。
物种多样性是评价群落稳定性的重要指标[23],可以反映一个群落的物种组成情况、复杂程度以及群落内物种丰富度等[15]。间作牧草后,苹果园杂草群落的多样性程度有所降低,各物种种群数量在群落中分布更随机,部分优势杂草在群落在中的优势度变高。间作牧草第1年,单播燕麦处理的物种多样性显著低于对照处理,同时群落优势种燕麦的优势度远高于其他物种,主要杂草如白草、牛膝菊、画眉草等物种重要值低于对照,说明种植燕麦后群落发生较大变化。试验第2年结果表明,鸭茅单播会降低群落物种丰富度、多样性与均匀度,并占据较高优势度,多年生牧草混播对于杂草群落多样性的影响同样显著。有研究表明,部分禾本科牧草在种植后会对杂草产生化感作用,影响杂草的生长[24],禾本科功能群叶面积和叶位相对较大,在生长方面占据优势,能抑制豆科、菊科杂草生长[25]。本研究中燕麦与鸭茅均为禾本科牧草,相较于豆科牧草,禾本科牧草处理中杂草群落物种多样性和丰富度较低,优势杂草重要值降低程度更大,这可能与禾本科牧草的上述特性有关。
“以草治草”是果园杂草生态防控的重要措施之一。栾好安[26]、张苗苗[27]通过对比植物群落中杂草密度、盖度、多度和重要值来确定杂草防除率,结果发现人工牧草有着良好的杂草防除效果。也有研究表明种植的牧草生物量越大,对杂草的抑制效果越好[28]。果园中种植人工牧草,会降低土壤中杂草种子密度,从而达到较好的杂草防控效果[29]。本研究中几种人工牧草均有着良好的杂草防除效果,但一年生和多年生牧草对杂草总相对盖度的影响程度受种植年限影响显著。间作牧草的第1年,种植一年生牧草的杂草防治效果良好,次年,种植多年生牧草的杂草防治效果更佳,这也与前面杂草群落重要值和多样性指数的变化一致。一年生和多年生牧草处理在种草第1、2年呈现出的这种相反的杂草防治效果,可能与所选牧草特性及土壤耕作有关:多年生牧草生长周期较长,试验第1年,植株仍处于苗期,根系较浅,生物量低,难以与原生的杂草竞争,杂草抑制效果也较差;试验第2年,多年生牧草已处于盛花期,生物量较大,对杂草的抑制效果逐渐体现。在土壤耕作方面,第1年试验地清耕对原生杂草影响较大,一年生牧草在清耕后播种,牧草生长周期短,能迅速占据空间,对杂草起到了较为明显的抑制效果;次年,牧草播种前仅对试验地进行浅耕,一年生杂草及深位性宿根杂草受影响较小,在牧草萌发前已占据一定生长优势,后续生长的牧草难以与之竞争。此外,牧草单、混播的种植模式也会对杂草群落产生影响,有研究表明混播牧草的杂草防治效果更佳[30],我们也得出了相似的结果,这可能是因为不同草种混杂一起生长,高低有序,相互穿插空隙,占据更多生长空间,使其他杂草生长的空间和光照竞争性减弱。
4. 结论
本研究分析了林芝河谷区苹果园间作不同牧草对果园内杂草群落多样性和组成的影响,得出以下主要结论:(1)林芝河谷区苹果园优势杂草种类主要有白草、画眉草、小头蓼、牛膝菊和蒲公英等;(2)间作牧草能改变杂草群落组成,减少杂草种类,降低杂草重要值;(3)不同间作牧草处理对杂草群落物种多样性影响有显著差异,但整体上降低了杂草群落的物种多样性,种植年限与种植模式对于杂草群落多样性和优势度均有着显著影响;(4)种植不同牧草种类对杂草均有一定的抑制效果,种植牧草的第1年一年生牧草的杂草抑制效果更好,第2年则为多年生牧草的杂草抑制效果更佳,禾本科牧草处理相较于豆科具有更好的杂草防控效果,混播处理的杂草防控效果优于单播。因此,本研究认为,针对果园的杂草生态防控,若果园会频繁翻耕,种植一年生禾本科牧草效果更好,若果园需长期种草,则种植多年生禾本科牧草或混播豆科、禾本科牧草效果更佳。
-
![]()
图 1 不同处理下杂草群落的聚类分析
虚线代表在该处对处理进行组别划分。处理简称同表1。下同。
Figure 1. Cluster analysis of weed communities in different treatments
Dashed line represents the grouping of treatments at this location. Treatments are the same as shown in Table 1. Same below.
![]()
图 2 不同处理下植物群落的Patrick指数与Simpson指数
不同小写字母表示不同间作牧草处理以及年限间的差异显著(P<0.05)。下同。
Figure 2. Patrick and Simpson indices of plant communities in different treatments
Different lowercase letters indicate significant differences at P<0.05 among different treatments in different years. Same below.
![]()
图 3 不同处理下植物群落Shannon指数与Pielou指数
Figure 3. Shannon and Pielou indices of plant communities in different treatments
表 1 不同处理牧草播种量
Table 1 Sowing of forages in different treatments
处理
Treatment简称
Abbreviation牧草品种
Forage species播种量
Seeding volume / g/m2一年生豆科单播 L1 箭筈豌豆Avena sativa L. 15 一年生禾本科单播 G1 燕麦Vicia sativa L. 30 一年生豆禾混播 L1G1 箭筈豌豆、燕麦 10.5、21 多年生豆科单播 L2 紫花苜蓿Medicago sativa L. 1.5 多年生禾本科单播 G2 鸭茅Dactylis glomerata L. 3 多年生豆禾混播 L2G2 紫花苜蓿、鸭茅 1.05、2.1 对照 CK − −  下载: 导出CSV
下载: 导出CSV
表 2 2021年不同处理中杂草群落组成及重要值
Table 2 Weed community composition and importance values in different treatments in 2021
科名
Family种名
Species处理 Treatment CK L1 G1 L1G1 L2 G2 L2G2 禾本科 画眉草 Eragrostis pilosa (L.) Beauv. 14.55 4.32 3.14 − 20.83 3.90 2.63 白草 Pennisetum flaccidum Griseb. 32.87 17.26 6.02 3.76 14.57 21.19 26.15 马唐 Digitaria sanguinalis (L.) Scop. − − − − − − 4.90 菊科 牛膝菊 Galinsoga parviflora Cav. 8.44 8.51 3.18 6.60 12.83 8.04 10.28 猪毛蒿 Artemisia scoparia Waldst. et Kit. 0.76 − − 0.80 − 0.77 − 蒲公英 Taraxacum mongolicum Hand.-Mazz. 4.75 3.08 6.27 2.66 6.16 5.12 2.50 秋鼠曲草 Pseudognaphalium hypoleucum(Candolle) Hilliard & B. L. Burtt 0.85 − − − − − 0.70 蓼科 小头蓼 Persicaria microcephala (D. Don) H. Gross 12.79 13.83 7.13 8.70 7.48 6.45 5.21 水蓼 Persicaria hydropiper (L.) Spach 0.95 − − − − 0.51 − 萹蓄 Polygonum aviculare L. 0.62 − 0.63 − 1.55 0.53 − 豆科 广布野豌豆 Vicia cracca L. − − − − − − 0.62 野苜蓿 Medicago falcata L. 5.96 3.24 2.99 0.62 1.41 8.16 0.74 白车轴草 Trifolium repens L. − − − − 4.59 − − 蔷薇科 绢毛委陵菜 Potentilla sericea L. 0.57 − − − 1.04 0.76 0.65 蛇莓 Duchesnea indica (Andr.) Focke − − − 0.69 − − 0.50 苋科 灰绿藜 Oxybasis glauca (L.) S. Fuentes, Uotila & Borsch 2.37 3.26 2.55 − 2.00 3.32 1.99 菊叶香藜 Dysphania schraderiana (Roemer & Schultes) Mosyakin & Clemants 0.76 − 1.37 2.64 4.78 1.43 1.92 莎草科 青绿薹草 Carex breviculmis R. Br. − 1.04 − − − − − 扁穗草 Blysmus compressus (L.) Panz. 0.63 − − − − 0.81 1.86 锦葵科 野葵 Malva verticillata L. 3.21 0.76 1.44 3.03 3.59 4.00 6.47 牻牛儿苗科 牻牛儿苗 Erodium stephanianum Willd. 1.17 − 0.64 − 2.10 − − 大麻科 大麻 Cannabis sativa L. 0.58 1.54 − − 2.48 0.53 0.48 天南星科 黄苞南星 Arisaema flavum (Forsk.) Schott 0.57 − − − − − − 灯芯草科 灯芯草 Juncus effusus L. 1.44 − − − − − 1.71 茜草科 拉拉藤 Galium spurium L. − − − − 0.49 − − 木贼科 节节草 Equisetum ramosissimum Desf. 1.30 − 0.63 0.63 − 0.52 3.48 注:处理简称同表1。下同。 Note: Treatments are the same as shown in Table 1. Same below.
下载: 导出CSV
表 3 2022年不同处理中杂草群落组成及重要值
Table 3 Weed community composition and importance values in different treatments in 2022
科名
Family种名
Species处理 Treatment CK L1 G1 L1G1 L2 G2 L2G2 禾本科 画眉草 Eragrostis pilosa (L.) Beauv. 15.06 10.08 3.46 11.89 10.21 1.83 3.19 白草 Pennisetum flaccidum Griseb. 34.58 54.87 15.46 27.84 6.69 − 4.88 菊科 牛膝菊 Galinsoga parviflora Cav. 7.43 6.49 10.94 21.08 1.33 1.72 1.50 蒲公英 Taraxacum mongolicum Hand.-Mazz. 10.01 10.87 15.24 5.19 13.45 7.26 3.04 秋鼠曲草 Pseudognaphalium hypoleucum (Candolle) Hilliard & B. L. Burtt 2.43 1.13 0.94 0.88 1.23 − − 一年蓬 Erigeron annuus (L.) Pers. 4.37 2.23 3.85 3.07 2.61 − − 白莲蒿 Artemisia stechmanniana Bess. 0.77 − − − 2.51 1.78 − 蓼科 尼泊尔酸模 Rumex nepalensis Spreng. 1.55 1.14 0.88 2.70 2.15 − − 小头蓼 Persicaria microcephala (D. Don) H. Gross 0.82 1.14 4.63 2.25 1.36 − − 萹蓄 Polygonum aviculare L. − 1.17 − − − − − 豆科 广布野豌豆 Vicia cracca L. 0.82 − 0.88 − − − − 野苜蓿 Medicago falcata L. 1.52 − − − − − − 蔷薇科 绢毛委陵菜 Potentilla sericea L. − − − 0.94 − 1.73 − 蛇莓 Duchesnea indica (Andr.) Focke − − 0.88 − − − − 唇形科 宝盖草 Lamium amplexicaule L. 2.64 1.13 2.11 1.90 1.51 − − 紫花香薷 Elsholtzia argyi Lévl. 2.37 − − − − − − 锦葵科 野葵 Malva verticillata L. 0.75 − − 0.91 − − 1.54 牻牛儿苗科 牻牛儿苗 Erodium stephanianum Willd. 2.89 2.28 3.76 1.22 − 3.36 2.99 大麻科 大麻 Cannabis sativa L. 2.99 1.18 1.01 − 1.27 − − 灯芯草科 灯芯草 Juncus effusus L. − − − − − 1.71 − 木贼科 节节草 Equisetum ramosissimum Desf. 2.16 − − − − − 1.64 车前科 毛车前 Plantago australis Lam. − − − − 1.26 − − 石竹科 鹅肠菜 Stellaria aquatica (L.) Scop. − 1.14 − − − − − 莎草科 青绿薹草 Carex breviculmis R. Br. 2.47 1.91 − − − − −
下载: 导出CSV
表 4 种植模式和种植年份与植物群落多样性指数的双因素方差检验
Table 4 Two-way ANOVA of treatment type and planting year on community diversity index of plant communities
影响因子
Impact factor因变量
Dependent
variable自由度
dfF值
F-valueP值
P-value年份 Patrick 1 14.313 0.001 Shannon 1 9.979 0.004 Pielou 1 1.816 0.189 Simpson 1 8.473 0.007 种植模式 Patrick 6 3.211 0.016 Shannon 6 5.254 0.001 Pielou 6 3.997 0.005 Simpson 6 4.335 0.003 年份 × 种植模式 Patrick 6 2.101 0.085 Shannon 6 7.713 0.000 Pielou 6 6.879 0.000 Simpson 6 8.419 0.000
下载: 导出CSV
-
[1] Gao QZ,Li Y,Xu HM,Wan YF,Jiang WZ. Adaptation strategies of climate variability impacts on alpine grassland ecosystems in Tibetan Plateau[J]. Mitig Adapt Strateg Glob Chang,2014,19(2):199−209. doi: 10.1007/s11027-012-9434-y
[2] 曹旭娟. 青藏高原草地退化及其对气候变化的响应[D]. 北京: 中国农业科学院, 2017: 5. [3] 余成群, 曲云鹤, 孙维, 武俊喜, 李少伟. 西藏“一江两河”地区饲草生产技术效率分析[C]//2015中国草原论坛论文集. 锡林浩特: 农业部草原监理中心, 中国草学会, 2015: 9-15. [4] 索朗曲吉,巴桑珠扎,旦增洛桑,李斌,次仁罗布,等. 西藏地区人工草地种植技术[J]. 畜牧兽医科学,2020(12):168−169. Suolangquji,Basangzhuzha,Danzengluosang,Li B,Cirenluobu,et al. Planting technology of artificial grassland in Tibet[J]. Graziery Veterinary Sciences,2020(12):168−169.
[5] 西藏自治区统计局, 国家统计局西藏调查总队. 西藏统计年鉴[M]. 北京: 中国统计出版社, 2021: 142-143. [6] 李晓刚,邵明灿,杨青松,王中华,蔺经. 梨园生草白三叶栽培对梨园杂草的抑制作用及其土壤理化性状的影响研究[J]. 上海农业科技,2017(2):106−107. [7] 陈清西,廖镜思,郑国华,刘殊. 果园生草对幼龄龙眼园土壤肥力和树体生长的影响[J]. 福建农业大学学报,1996,25(4):429−432. Chen QX,Liao JS,Zheng GH,Liu S. Effects of sown grass mulch on soil fertility and tree growth of young Longan orchard[J]. Journal of Fujian Agricultural University,1996,25(4):429−432.
[8] 王依,陈成,马拦妮,雷靖,徐明,雷玉山. 行间生草对秦岭北麓猕猴桃园土壤养分、pH值的影响[J]. 中国农学通报,2019,35(15):59−65. Wang Y,Chen C,Ma LN,Lei J,Xu M,Lei YS. Inter-row grass:effects on soil nutrients and pH value of kiwifruit orchards in northern Qinling mountains[J]. Chinese Agricultural Science Bulletin,2019,35(15):59−65.
[9] 韦红霞,王妍,郗小燕,崔连超. 果园生草对苹果产量及果实品质的影响[J]. 果农之友,2022(10):4−5. doi: 10.3969/j.issn.1671-7759.2022.10.002 [10] Sawicka B,Krochmal-Marczak B,Barbaś P,Pszczółkowski P,Ćwintal M. Biodiversity of weeds in fields of grain in south-eastern Poland[J]. Agriculture,2020,10(12):589. doi: 10.3390/agriculture10120589
[11] 冉海燕,兰献敏,陈仕红,叶照春. 果园以草治草研究进展[J]. 农技服务,2019,36(2):68. [12] 肖润林,向佐湘,徐华勤,单武雄,陈佩,等. 间种白三叶草和稻草覆盖控制丘陵茶园杂草效果[J]. 农业工程学报,2008,24(11):183−187. Xiao RL,Xiang ZX,Xu HQ,Shan WX,Chen P,et al. Ecological effects of the weed community in tea garden with intercropping white clover and straw mulching[J]. Transactions of the Chinese Society of Agricultural Engineering,2008,24(11):183−187.
[13] 王加国,李苇洁,吴迪,韩振诚,苏文文. 鼠茅草对猕猴桃果园抑草效果及土壤肥力的影响[J]. 中国南方果树,2022,51(4):109−113. [14] 王晶晶,李娜,张身嗣. 覆盖作物白三叶对蓝莓园杂草的生物防除效果[J]. 北方园艺,2017(3):138−140. Wang JJ,Li N,Zhang SS. Biological control of cover crops white clover against weeds in blueberry orchard[J]. Northern Horticulture,2017(3):138−140.
[15] 王雪,张威,伍玉鹏,胡荣桂,姜炎彬. 绿肥种植配施减量氮肥对柑橘园杂草群落组成及多样性的影响[J]. 植物科学学报,2020,38(2):212−220. Wang X,Zhang W,Wu YP,Hu RG,Jiang YB. Effects of natural grasses and intercropping green manure on the composition and diversity of the weed community in a citrus orchard[J]. Plant Science Journal,2020,38(2):212−220.
[16] 张莉. 耕作方式对华北寒旱区农田杂草群落结构及土壤物理特性的影响[D]. 保定: 河北农业大学, 2014: 4. [17] 包赛很那,苗彦军,王向涛,赵玉红,孙磊,王明涛. 开花期黄花草木樨根浸提液对林芝市4种主要农田杂草的化感作用[J]. 高原农业,2020,4(4):380−385. Baosaihenna,Miao YJ,Wang XT,Zhao YH,Sun L,Wang MT. Allelopathic effect of extract of melilotus officinalis root in flowering stage on four main farmland weeds in Nyingchi region[J]. Journal of Plateau Agriculture,2020,4(4):380−385.
[18] 郭金丽,李青丰. 克隆整合对白草克隆生长的影响[J]. 中国草地学报,2008,30(6):43−48. Guo JL,Li QF. Effects of clonal integration on clonal growth in Pennisetum centrasiaticum[J]. Chinese Journal of Grassland,2008,30(6):43−48.
[19] 郭丽珠,黄顶,张丛,李佳欢,赵欢,王堃. 退化典型草原狼毒生物量分配及异速生长分析[J]. 中国草地学报,2019,41(6):53−59. Guo LZ,Huang D,Zhang C,Li JH,Zhao H,Wang K. Analysis of biomass allocation and allometric growth of Stellera chamaejasme in degraded typical steppe[J]. Chinese Journal of Grassland,2019,41(6):53−59.
[20] 张鲜花,朱进忠,李海琪. 天山北坡东段与西段不同海拔鸭茅群落特征及物种多样性研究[J]. 草地学报,2016,24(4):760−767. Zhang XH,Zhu JZ,Li HQ. Species diversity and characteristics of Dactylis glomerata community in different altitudes on the eastern and western sections of the northern slope of Tianshan mountain[J]. Acta Agrestia Sinica,2016,24(4):760−767.
[21] 朱先波,潘亮,王华玲,肖丽丽,钟云鹏,等. 十堰猕猴桃果园生草生态效应的分析[J]. 农业资源与环境学报,2020,37(3):381−388. Zhu XB,Pan L,Wang HL,Xiao LL,Zhong YP,et al. Analysis of the ecological effects of grass-growing in kiwifruit orchards in Shiyan,China[J]. Journal of Agricultural Resources and Environment,2020,37(3):381−388.
[22] 刘芳,任启飞,马菁华,欧明烛,陈云飞. 入侵植物牛膝菊研究进展[J]. 农业与技术,2022,42(14):34−37. [23] Tilman D,Isbell F. Biodiversity recovery as nitrogen declines[J]. Nature,2015,528(7582):336−337. doi: 10.1038/nature16320
[24] Cornelius CD,Bradley KW. Herbicide programs for the termination of various cover crop species[J]. Weed Technology,2017,31(4):514−522. doi: 10.1017/wet.2017.20
[25] 张力天,杨时海,刘炜,王晓丽,秦金萍,等. 雅鲁藏布江中上游白草主要分布区植物群落特征[J]. 生态学报,2022,42(15):6111−6124. Zhang LT,Yang SH,Liu W,Wang XL,Qin JP,et al. Characteristics of plant communities in distribution areas of Pennisetum centrasiaticum in the upper and middle reaches of the Yarlung Zangbo River[J]. Acta Ecologica Sinica,2022,42(15):6111−6124.
[26] 栾好安. 三峡库区橘园绿肥的生态效应及其对柑橘产量和品质的影响[D]. 武汉: 华中农业大学, 2015: 2. [27] 张苗苗. 果园生草对杂草及土壤微生物的影响[D]. 海口: 海南大学, 2020: 7. [28] Latif S,Gurusinghe S,Weston PA,Brown WB,Quinn JC,et al. Performance and weed-suppressive potential of selected pasture legumes against annual weeds in south-eastern Australia[J]. Crop & Pasture Science,2019,70(2):147−158.
[29] Scavo A,Restuccia A,Abbate C,Lombardo S,Fontanazza S,et al. Trifolium subterraneum cover cropping enhances soil fertility and weed seedbank dynamics in a Mediterranean apricot orchard[J]. Agronomy for Sustainable Development,2021,41(6):70. doi: 10.1007/s13593-021-00721-z
[30] 孙瑞红,姜莉莉,宫庆涛,武海斌,蒋恩顺. 落叶果园混合生草种植管理与作用效果[J]. 特种经济动植物,2020,23(9):45−47.




计量
- 文章访问数: 283
- HTML全文浏览量: 43
- PDF下载量: 149
