
Response of osmoregulatory substances and antioxidant enzyme activities in heteromorphic leaves of Populus euphratica Oliv. to groundwater depth
-
摘要:
以塔里木荒漠区建群种胡杨(Populus euphratica Oliv.)为实验材料,研究3种典型异形叶的渗透调节物质和抗氧化酶活性沿地下水埋深(GWD)梯度的变化规律及适应干旱胁迫的策略。结果显示:(1)随GWD的增加,3种异形叶的相对含水量、叶水势、可溶性蛋白质(SP)含量和过氧化物酶(POD)活性呈降低趋势,水分饱和亏缺、游离脯氨酸(Pro)含量、可溶性糖(SS)含量和丙二醛(MDA)含量呈增加趋势,而超氧化物歧化酶(SOD)始终保持较高活性且较稳定。(2)条形叶Pro、SS含量与GWD呈极显著正相关,卵形叶SS、SP含量与GWD呈显著正相关,锯齿阔卵形叶SS含量与GWD呈极显著正相关。(3)条形叶主要以Pro和SS协同维持细胞渗透势以应对水分胁迫,卵形叶以增强渗透调节能力(SP、Pro、SS)来提高其抗旱性,锯齿阔卵形叶则以SS、Pro维持细胞膨压,并通过POD、SOD清除活性氧(ROS)协同适应干旱的荒漠环境。
Abstract:Populus euphratica Oliv., an established species in the Tarim Desert Area, was used to investigate variations in osmoregulatory substances and antioxidant enzyme activities of three typical heteromorphic leaves along the groundwater depth (GWD) gradient as well as differences in their adaptive strategies to drought stress. Results showed that: (1) with the increase in GWD, the relative water content, leaf water potential, soluble protein (SP) content, and peroxidase (POD) activity of the three heteromorphic leaves of P. euphratica showed a decreasing trend, while the water saturation deficit, free proline (Pro) content, soluble sugar (SS) content, and malondialdehyde (MDA) content showed an increasing trend, and superoxide dismutase (SOD) maintained high and stable activity. (2) The Pro and SS contents in lanceolate leaves showed a highly significant positive correlation with GWD, the SS and SP contents in oval leaves showed a significant positive correlation with GWD, and the SS content in serrated broad-oval leaves showed a highly significant positive correlation with GWD. (3) The lanceolate leaves maintained cellular osmotic potential through the synergistic action of Pro and SS to cope with water stress, oval leaves enhanced drought resistance through osmotic adjustment (SP, Pro, SS), and serrated broad-oval leaves maintained cellular expansion pressure with SS and Pro, while eliminating reactive oxygen species (ROS) through POD and SOD, which synergistically adapted to increasingly arid desert environments.
-
在全球气候变化大背景下,极端天气事件频发、大气环流与水碳循环格局变化等导致的干旱强度、频率和持续时间不断增加,进而造成全球范围内森林大面积衰退、死亡事件逐年激增,植物对干旱胁迫的响应与适应已经成为植物生理生态学研究的热点[1, 2]。我国是干旱灾害发生频率较高、影响也较严重的国家之一,尤其是我国四大生态脆弱区之一的塔里木盆地,降水稀少,地表水匮乏,地下水成为荒漠植被赖以生存的重要水分来源。近年来随着全球变暖及区域水资源的过度开采,地下水埋深(GWD)持续降低,直接影响着植物水分和养分吸收、供应及各种生理生化代谢过程,威胁荒漠植被的生存发展与区域生态安全[3-5]。
极端干旱荒漠生境中,植物体内活性氧(ROS)代谢的动态平衡被打破[6],生物膜结构与功能被破坏,植物积累大量渗透调节物质并提高抗氧化酶活性,借此来稳定原生质胶体和生物膜以及组织内的代谢过程,以应对长期干旱缺水的环境[7-10]。Wu等[11]研究表明,随GWD下降,黎明前的梭梭(Haloxylon ammodendron (C. A. Mey.) Bunge)叶片水势和碳同化作用显著降低。干旱胁迫显著降低大山樱(Prunus sargentii Rehder)的叶水势和蒸腾作用,而日本落叶松(Larix kaempferi (Lamb.) Carr.)变化不明显,表现出更大的抗性和弹性[12]。沙芥(Pugionium cornutum (L.) Gaertn.)和斧形沙芥(P. dolabratum Maxim.)叶中丙二醛(MDA)含量随干旱胁迫程度加剧呈升高趋势,过氧化物酶(POD)活性先升高后降低,超氧化物歧化酶(SOD)活性先降低后升高[13]。郭艳阳等[14]研究发现玉米(Zea mays L.)叶片MDA含量随水分胁迫的加剧持续增加,SOD、POD活性先升高后降低。
胡杨(Populus euphratica Oliv.)为杨柳科杨属植物,是荒漠河岸林的建群种和荒漠地区特有的珍贵树种[15]。胡杨具有典型的异形叶性,在幼苗、幼树和成年树的下部着生条形或披针形叶,在成年树中部开始出现过渡形叶如卵形或肾形叶,而成年树上部则着生成熟的锯齿卵圆形或锯齿阔卵形叶[16, 17]。生长在河岸沙坡上的同龄树,其条形叶明显少于河岸低地[18];具有叶齿的叶片在干旱胁迫条件下存在明显优势[19],且锯齿叶趋于旱生性结构[20],对干旱环境的适应能力更强[21, 22]。因此,随着地下水位的下降,可能出现条形叶逐渐减少以至消失,而锯齿阔卵形叶逐渐占据树冠上层空间的现象。前人对胡杨异形叶光合、水分生理及其相关基因开展了较多研究[23-25],但关于异形叶渗透调节与保护酶对GWD的响应格局及其适应策略鲜见报道。
本研究以塔里木极端干旱区的胡杨为材料,在测定地下水埋深梯度下3种典型异形叶(条形、卵形、锯齿阔卵形)的水分生理与抗逆物质的基础上,试图回答以下问题:(1)3种异形叶水力性状及抗逆生理物质沿GWD梯度的变异格局及差异如何?(2)3种异形叶应对荒漠环境的生理适应策略是否存在差异?(3)从抗逆生理的角度上,如何解释随GWD的增加,条形叶逐渐消失、锯齿阔卵形叶占据树冠空间?研究结果旨在揭示胡杨3种异形叶适应极端干旱区日益旱化荒漠环境的生理生态对策,阐明叶形变化的原因,为极端干旱区胡杨林的保育复壮及荒漠河岸林生态恢复提供科学依据。
1. 材料与方法
1.1 研究区概况
研究区位于新疆塔里木河上游,海拔800~1 300 m,年均太阳辐射133.7~146.3 kcal/cm2,年均日照2 556.3~3 031.2 h,年均气温10.6 ℃,年均日较差15 ℃,年平均≥10 ℃稳定积温达3953 ℃,无霜期190~251 d,年均降水量40.1~82.5 mm,年均蒸发量1 876.6~2 558.9 mm,年平均风速2.3 m/s,多为西北风或西风。2022年生长季于阿瓦提县丰收三场、塔河源、南口镇、沙雅县天然胡杨林内,垂直河岸设置50 m宽的样带,根据植物群落结构组成、胡杨密度、长势情况设置3~5个50 m×50 m的调查样方。采用机械打井,建立地下水位观测井(PVC管,10 m),用于长期监测GWD的变化。GWD采用电导法测定[26],各样地GWD见表1。
表 1 样地概况Table 1. Characteristics of study plots样地
Site经纬度
Longitude and latitude地下水埋深
GWD / m密度
Density / ind./hm2平均树龄
Average age / a平均树高
Average height / m平均胸径
Average DBH / cm塔河源 40°30′04″N, 80°58′27″E 1.0~1.5 290.5 80.2 6.4 26.3 南口镇 40°26′29″N, 81°09′10″E 2.3~2.7 171.3 69.1 8.5 22.6 阿瓦提 40°19′44″N, 80°23′35″E 4.0~4.5 268.1 64.6 7.9 21.1 沙雅县 40°41′19″N, 81°59′43″E 7.5~8.0 120.2 110.6 7.8 36.4 1.2 样品采集
2022年7-8月,在塔河源、南口镇、阿瓦提、沙雅县4个GWD样地,分别随机选择健康、无病虫害、叶形分化明显的成熟胡杨树10株进行采样,在树冠上、中、下层3个高度四周方向剪下生长良好的当年生小枝(末端无分枝),每株采集条形、卵形、锯齿阔卵形叶(图1)各3~5片,用湿润纱布包好,装入自封袋,置于冷藏箱中带回实验室,一部分用于测定叶片水分状况,另一部分放入−80 ℃冰箱保存。
![]() 图 1 胡杨3种异形叶特征A:条形叶;B:卵形叶;C:锯齿阔卵形叶。Figure 1. Characteristics of three heteromorphic leaves of Populus euphraticaA: Lanceolate leaf; B: Oval leaf; C: Serrated broad-oval leaf.
图 1 胡杨3种异形叶特征A:条形叶;B:卵形叶;C:锯齿阔卵形叶。Figure 1. Characteristics of three heteromorphic leaves of Populus euphraticaA: Lanceolate leaf; B: Oval leaf; C: Serrated broad-oval leaf.1.3 测定指标与方法
1.3.1 叶水势测定
野外采用PSYPRO露点水势仪(Wescor,美国)测定叶片水势(Ψleaf),选取成熟健康的胡杨3种异形叶,将叶片中部置于叶室(L-51)中,拧紧L-51探测头,并用密封条将叶室周围空隙密封,平衡30 min后进行测定[27],每株每叶形重复3次。
1.3.2 水分生理参数测定
用精度为0.000 1 g的电子天平称量叶片鲜重(Wf),每株每叶形重复3次,然后将叶片浸入蒸馏水中12 h以上,直至叶片吸水饱和,称量叶片饱水重(Wt),再将叶片放置于烘箱中105 ℃杀青30 min、80 ℃烘干至恒重,称量叶片干重(Wd),根据公式计算叶片相对含水量,RWC=(Wf−Wd)/(Wt−Wd)×100%,叶片水分饱和亏缺WSD=(Wt−Wf)/(Wt−Wd)×100%[24, 28]。
1.3.3 抗逆生理指标测定
将3种异形叶分别混合取样,各生理指标测定方法均参考《植物生理学实验指导》[29],所有测定重复3次。
1.3.3.1 渗透调节物质测定
游离脯氨酸(Pro)含量采用磺基水杨酸提取、酸性茚三酮法测定;可溶性糖(SS)含量采用硫酸-蒽酮比色法测定;可溶性蛋白(SP)含量采用考马斯亮蓝G-250染色法测定。
1.3.3.2 MDA含量及抗氧化酶活性测定
酶液提取:称取混样2.0 g,放入预先冰冻的研钵中,加入少量石英砂和0.05 mol/L pH值为7.8的磷酸缓冲液(PBS),冰浴研磨成匀浆,转移至50 mL容量瓶准确定容。定容后的溶液4 ℃、10 000 r/min离心15 min,上清液即提取液,用于MDA、SOD和POD测定。
MDA含量采用硫代巴比妥酸显色法测定, SOD活性采用氮兰四唑(NBT)光化还原法测定,以抑制NBT还原率50%为1个酶活单位(U),POD活性采用愈创木酚氧化法测定,以1 min内A470变化0.1为1个酶活单位(U)。
1.4 数据分析
使用Excel 2019软件对原始数据进行整理,运用SPSS统计软件对不同GWD同种叶形、同一GWD不同叶形的生理指标进行单因素方差分析(One-way ANOVA)与Duncan多重比较。对各生理指标之间的关系及其与GWD之间的关系进行Pearson相关分析,对3种异形叶生理指标沿GWD梯度的协变关系进行主成分分析(PCA)。利用Origin 2021及R 4.2.3软件进行绘图。
2. 结果与分析
2.1 胡杨异形叶水分生理参数沿GWD的变化
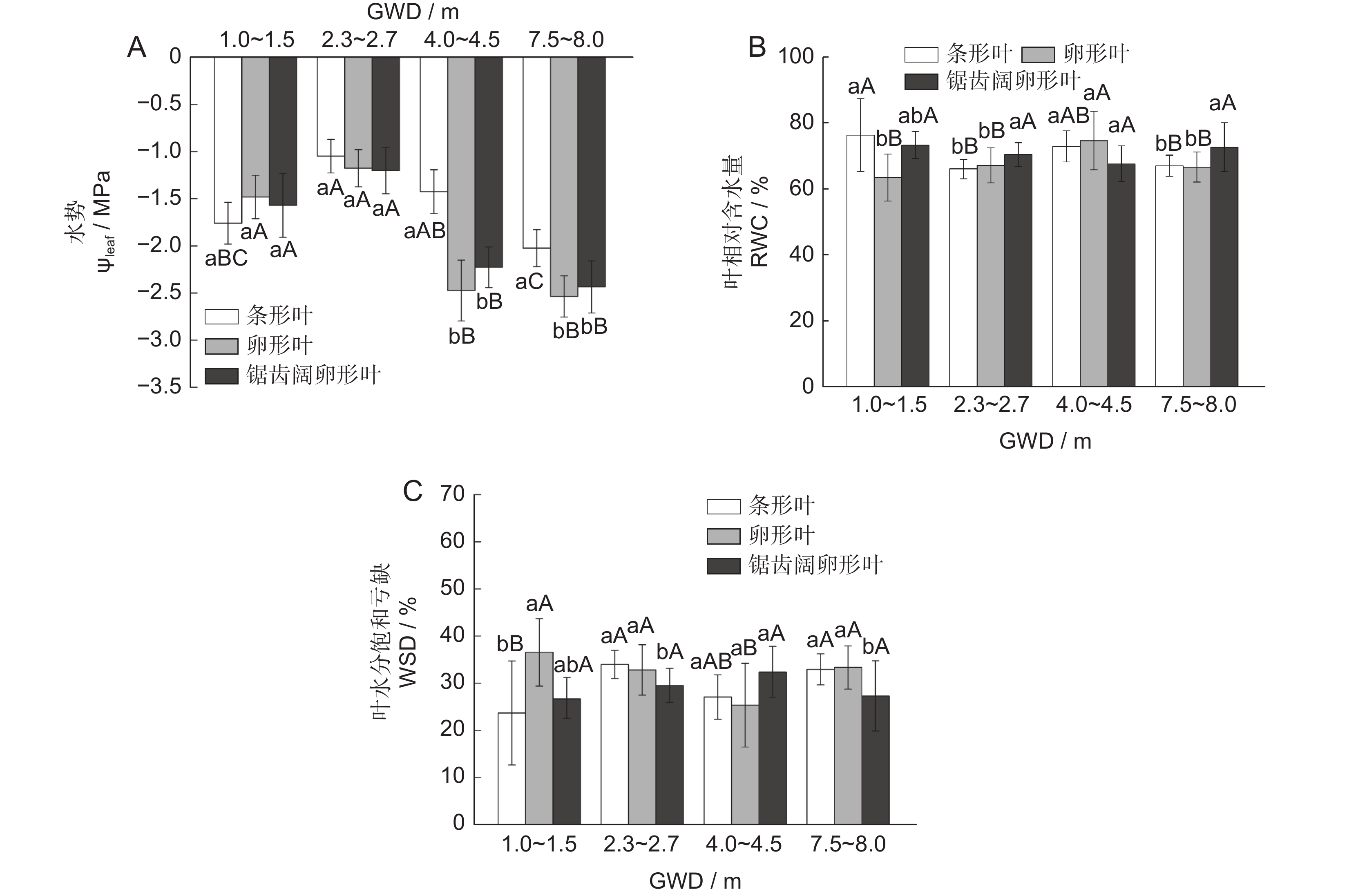
随着GWD的增加,胡杨异形叶水势日均值、RWC呈下降趋势(图2:A、B),WSD呈升高趋势(图2:C)。GWD在 7.5~8.0 m时,3种异形叶的水势日均值最低,与GWD 2.3~2.7 m时的同种叶形相比,条形、卵形、锯齿阔卵形叶分别下降了1.93、2.16、2.03倍。方差分析表明,不同GWD生境下的胡杨3种异形叶水势日均值差异显著(P<0.05),条形叶与卵形叶的RWC、WSD差异显著。同一GWD生境,GWD>3.0 m时条形叶水势日均值与其他两种叶形差异显著,WSD仅在GWD 4.0~4.5 m时3种叶形间有显著差异,表明随GWD增加,胡杨异形叶所受干旱胁迫的程度逐渐加剧,锯齿阔卵形叶维持叶片水分稳定的能力强于其他两种叶形。
![]() 图 2 异形叶水势、相对含水量、水分饱和亏缺对地下水埋深梯度的响应不同小写字母表示同一样地不同叶形间差异显著(P<0.05),不同大写字母表示同一叶形不同样地间差异显著(P<0.05);GWD:地下水埋深。下同。Figure 2. Response of water potential, relative water content, and water saturation deficit in heteromorphic leaves to GWDDifferent lowercase letters indicate significant differences among different leaf shapes in the same site (P<0.05), and different uppercase letters indicate significant differences among different sites in the same leaf (P<0.05); GWD: Groundwater depth. Same below.
图 2 异形叶水势、相对含水量、水分饱和亏缺对地下水埋深梯度的响应不同小写字母表示同一样地不同叶形间差异显著(P<0.05),不同大写字母表示同一叶形不同样地间差异显著(P<0.05);GWD:地下水埋深。下同。Figure 2. Response of water potential, relative water content, and water saturation deficit in heteromorphic leaves to GWDDifferent lowercase letters indicate significant differences among different leaf shapes in the same site (P<0.05), and different uppercase letters indicate significant differences among different sites in the same leaf (P<0.05); GWD: Groundwater depth. Same below.2.2 胡杨异形叶渗透调节物质含量沿GWD的变化
随GWD的增加,胡杨异形叶Pro、SS含量呈升高趋势(图3:A、B),SP含量呈下降趋势(图3:C)。GWD在 4.0~4.5 m时,3种异形叶的Pro含量均达最高值,与GWD在 1~1.5 m时的同种叶形相比,条形、卵形与锯齿阔卵形叶分别增加了1.83、3.90、2.80倍。方差分析表明,不同GWD生境下胡杨3种异形叶Pro、SS及SP含量的差异均达显著水平。同一GWD生境,条形叶与锯齿阔卵形叶的Pro、SS含量差异显著;除GWD 7.5~8.0 m外,不同生境3种叶形间的SP含量差异均达显著水平,且随GWD增加,锯齿阔卵形叶SP含量均显著高于其他两种叶形。这表明胡杨3种异形叶对GWD梯度的响应存在差异,其中锯齿阔卵形叶的SP含量受GWD变化的影响最大。
![]() 图 3 异形叶渗透调节物质(Pro、SS、SP)含量对地下水埋深梯度的响应Figure 3. Response of osmoregulatory substances (Pro, SS, SP) in heteromorphic leaves to GWD
图 3 异形叶渗透调节物质(Pro、SS、SP)含量对地下水埋深梯度的响应Figure 3. Response of osmoregulatory substances (Pro, SS, SP) in heteromorphic leaves to GWD2.3 异形叶MDA含量及抗氧化酶活性沿GWD的变化
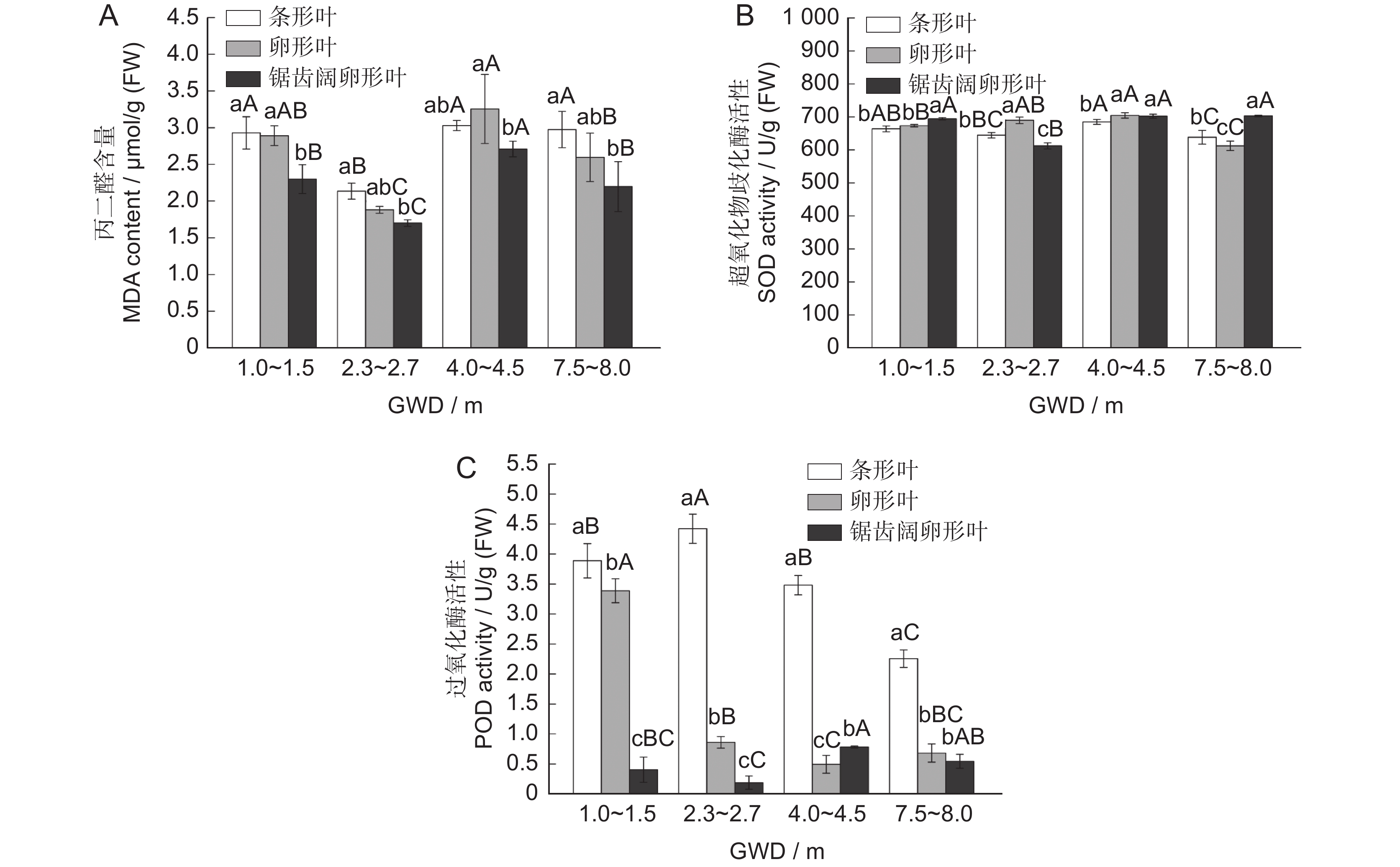
随GWD的增加,胡杨异形叶的MDA含量呈升高趋势(图4:A),SOD一直维持较高活性水平(图4:B),POD活性呈下降趋势(图4:C)。GWD在 4.0~4.5 m时,3种异形叶的MDA含量积累最多,且SOD活性达最高值。与GWD 为1~1.5 m时相比,GWD在 7.5~8.0 m时,条形叶、卵形叶的POD活性分别下降41.99%、79.84%。方差分析表明,不同GWD生境下胡杨3种异形叶的MDA含量、SOD及POD活性差异显著。同一GWD生境下,条形叶的MDA含量高于其他两种叶形,且差异显著,SOD活性在条形叶与锯齿阔卵形叶间差异显著;除GWD 7.5~8.0 m外,POD活性在3种叶形间差异显著,其中条形叶的POD活性始终保持较高水平,平均比卵形叶、锯齿阔卵形叶高4.16、10.40倍。这表明随着GWD的增加与干旱胁迫程度的加剧,条形叶膜脂过氧化程度明显高于其他两种叶形。3种异形叶通过提高SOD、POD的活性,减轻膜损伤,增强植物的抗旱性。
![]() 图 4 异形叶MDA含量及SOD、POD活性对地下水埋深梯度的响应Figure 4. Response of MDA content and SOD and POD activities in heteromorphic leaves to GWD
图 4 异形叶MDA含量及SOD、POD活性对地下水埋深梯度的响应Figure 4. Response of MDA content and SOD and POD activities in heteromorphic leaves to GWD2.4 胡杨异形叶生理指标与地下水埋深的相关性
Pearson相关性分析结果表明,条形叶Pro、SS与GWD呈极显著正相关,POD与GWD呈极显著负相关(表2)。卵形叶SS、SP与GWD呈显著正相关,POD与GWD呈极显著负相关(表3)。锯齿阔卵形叶SS与GWD呈极显著正相关(表4)。说明在干旱胁迫下,胡杨3种异形叶的生理指标与GWD变化密切相关,渗透调节物质的积累、抗氧化酶活性的变化与膜脂过氧化几乎同步。
表 2 条形叶生理指标与GWD的相关关系矩阵Table 2. Correlation matrix of GWD and six physiological indicators of lanceolate leaves指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.772** 0.709** −0.360 0.291 −0.194 −0.798** Pro 1.000 0.585* −0.379 0.733** 0.340 −0.754** SS 1.000 0.339 0.519 −0.299 −0.929** SP 1.000 0.119 −0.425 −0.159 MDA 1.000 0.440 −0.649* SOD 1.000 0.149 POD 1.000 注:*表示相关性显著(P<0.05),**表示相关性极显著(P<0.01)。下同。 Notes: *, significant (P<0.05); **, highly significant (P<0.01). Same below. 表 3 卵形叶生理指标与GWD的相关关系矩阵Table 3. Correlation matrix of GWD and six physiological indicators of oval leaves指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.441 0.617* 0.667* 0.098 −0.523 −0.799** Pro 1.000 0.342 0.801** 0.618* 0.395 −0.528 SS 1.000 0.760** 0.639* −0.628* −0.117 SP 1.000 0.720** −0.146 −0.389 MDA 1.000 0.082 0.174 SOD 1.000 0.034 POD 1.000 表 4 锯齿阔卵形叶生理指标与GWD的相关关系矩阵Table 4. Correlation matrix of GWD and six physiological indicators of serrated broad-oval leaves指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.392 0.840** 0.318 0.198 0.337 0.468 Pro 1.000 0.789** 0.188 0.707* 0.402 0.762** SS 1.000 0.122 0.621* 0.599* 0.784** SP 1.000 −0.430 −0.708* −0.216 MDA 1.000 0.760** 0.738** SOD 1.000 0.742** POD 1.000 2.5 胡杨异形叶生理指标的主成分分析
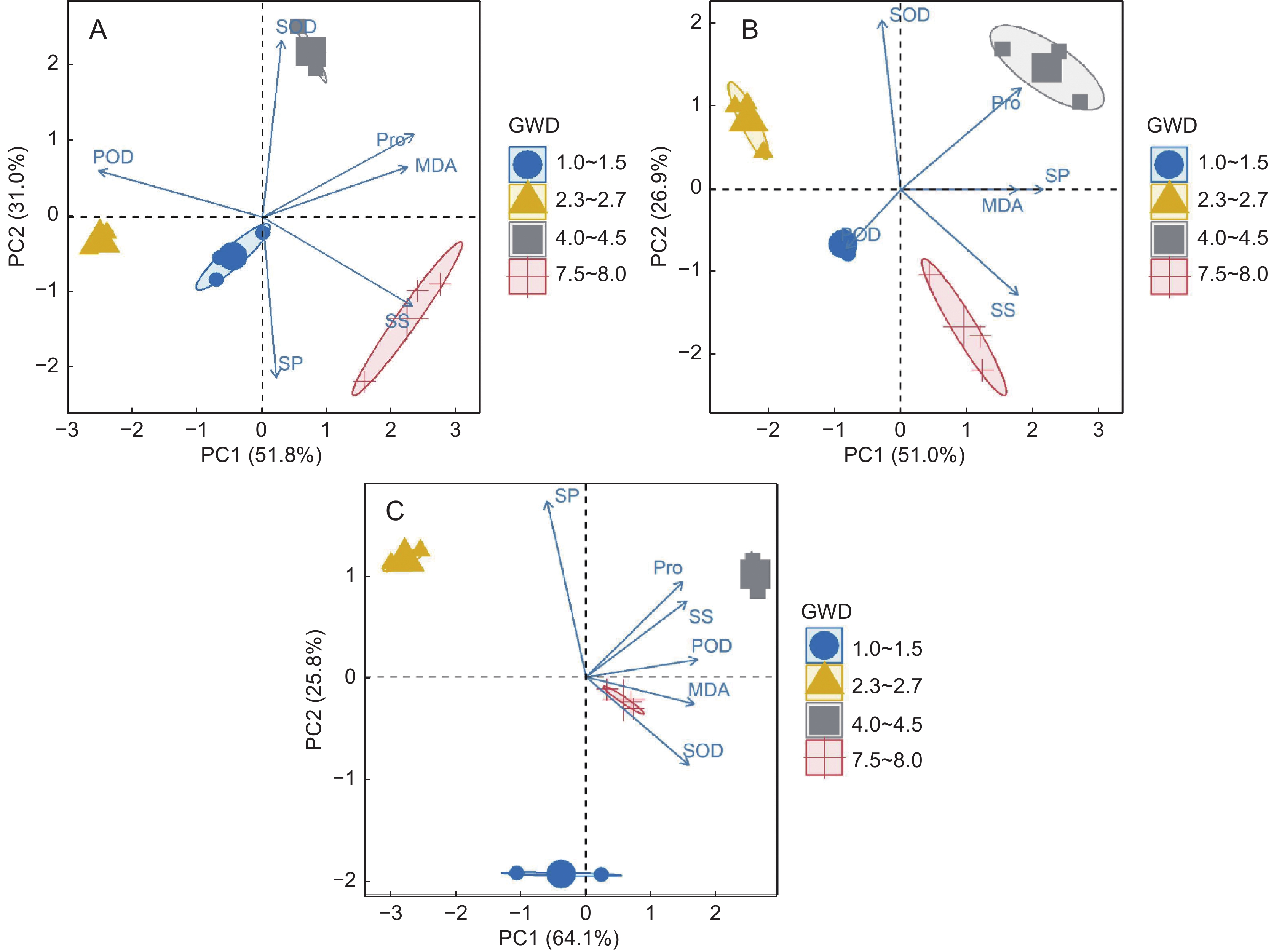
本研究对胡杨3种异形叶的生理指标进行了主成分分析。结果发现,条形叶前2个特征根累计贡献率达82.8 %(图5:A),PC1从左至右GWD逐渐增加,Pro、SS和MDA特征值均较高,且与PC1正相关。卵形叶第Ⅰ主成分贡献率为51.0 %,第Ⅱ主成分贡献率为26.9 %(图5:B),SP、Pro以及SS在PC1上均有较大的正载荷,其中SP与MDA有强烈的正相关,SOD在PC2上有最大的正载荷。锯齿阔卵形叶前2个特征根累计贡献率达89.9 %(图5:C),第Ⅰ主成分贡献率为64.1 %,为主要综合因子,除SP与其相关性较弱外,其他指标均与PC1密切正相关,第Ⅱ主成分中SP的特征值最高。
![]() 图 5 条形叶(A)、卵形叶(B)、锯齿阔卵形叶(C)生理指标的主成分分析Figure 5. Principal component analysis of physiological traits of lanceolate (A), oval (B), and serrated broad-oval leaves (C)
图 5 条形叶(A)、卵形叶(B)、锯齿阔卵形叶(C)生理指标的主成分分析Figure 5. Principal component analysis of physiological traits of lanceolate (A), oval (B), and serrated broad-oval leaves (C)3. 讨论
极端干旱荒漠区降水稀少,GWD是决定植物生存、生长和发育状况的关键因素[30]。植物水势能够反映其所受干旱胁迫的程度[31],水势越低,表明受干旱胁迫的程度越高。RWC表征植物组织的水分状况,WSD愈大,植物组织水分亏缺愈严重[24, 28]。本研究表明,随GWD增加引起水分运输距离的逐渐增加[17],3种异形叶的水力性状发生不同程度的变化,叶片中的淀粉向SS转化,以降低植物水势,增加土壤与植物之间的水势差,从而增强蒸腾拉力来保证水分的正常运输,满足自身生命活动的水分需求[32]。
植物抗逆性与生物膜结构及其功能的稳定性密切相关。干旱胁迫可对细胞膜造成伤害,导致渗透物质的大量外流,引起电导率上升[33]。Pro、SS和SP对土壤水分的变化非常敏感[34],在植物遭受干旱胁迫时能够降低细胞渗透势,其积累量常被作为植物抗旱性强弱的标志[35]。本研究发现,胡杨通过代谢途径的改变来增加Pro、SS的含量,从而维持一定的细胞渗透势以抵御干旱胁迫;而SP合成受阻,可能是由于干旱胁迫下,膜脂过氧化作用加剧、细胞膜受损,同时叶片光合碳同化效率下降等原因共同导致,这与韩蕊莲等[36]的研究结果一致。
植物在逆境下遭受伤害或器官衰老时,膜脂过氧化产物MDA的积累增多,从而对植物代谢造成严重干扰,引起一系列生理生化变化[37]。MDA含量的变化一定程度上可反映脂膜的损伤程度[38]。本研究结果表明,干旱胁迫下,条形叶和卵形叶的脂膜受伤害程度高于锯齿阔卵形叶,细胞结构破坏严重。锯齿阔卵形叶的脂膜损伤程度最轻,表明其抗旱能力高于条形叶和卵形叶,这与李萍萍等[39]的结果相一致。
SOD和POD是植物体内清除ROS的重要细胞保护酶类,其活性高低可以反映植物对逆境胁迫的适应能力[40]。本研究结果显示,SOD在3种异形叶的抗氧化酶防御系统中均发挥重要作用;而POD活性呈下降趋势,可能是由于氧化胁迫导致的脂膜损伤程度严重,且SP的合成能力下降,超出了POD的调控范围[41]。POD相较于SOD对GWD的变化更为敏感,并在条形叶适应干旱胁迫的策略中发挥作用。SOD和POD协同调节以清除ROS,减轻干旱胁迫引发的伤害,这与李端等[42]的研究结果一致。
胡杨作为生长在干旱荒漠地区的特有树种,正是以其异形叶形态解剖结构、抗氧化等生理生化特性的差异来适应极端干旱的生长环境[39]。生理指标与GWD的相关性分析结果表明,渗透调节物质和抗氧化酶均在胡杨异形叶对抗逆境过程中发挥重要作用,但3种异形叶的抗旱策略不同。随GWD的增加,条形叶主要通过积累大量的Pro和SS进行渗透调节来发挥作用,从而增强细胞的抗脱水能力;卵形叶则通过SP、Pro、SS 3种渗透调节物质共同调节渗透势,从而适应外界环境的变化;而锯齿阔卵形叶则迅速提高POD和SOD的活性,同时加快光合速率[43]积累大量的SS,与Pro一起发挥渗透调节作用,使植物体维持正常代谢水平。
综上所述,GWD是影响胡杨异形叶水力及生理性状的关键限制因子,3种异形叶通过形态结构和生理性状的变化,以及性状之间协同-权衡的生态对策来适应极端干旱的荒漠环境。随着全球变暖与干旱区GWD的持续增加,锯齿阔卵形叶作为最趋成熟的一种叶形,能够充分利用渗透调节物质和抗氧化酶两大系统协同减轻干旱胁迫的伤害,抗旱能力更强,从而使其能够在极端干旱的荒漠环境下得以生存,并占据树冠的主体,这可能是胡杨叶形随生存环境而变化的原因。
4. 结论
胡杨异形叶水分与抗逆生理指标对GWD变化具有明显的响应特征,但3种异形叶对干旱逆境的适应策略存在明显差异。随GWD的增加,从土壤至胡杨枝叶的水分运输距离增加,水分向上运输的阻力与水力限制增强,锯齿阔卵形叶通过渗透调节物质和抗氧化酶两大系统的协同作用来降低水势,维持细胞膜和组织水分的相对稳定。因此,抗旱性弱的条形叶大多分布在树冠下层,并随GWD的增加逐渐消失,而抗旱性强的锯齿阔卵形叶逐渐占据树冠上层空间。
-
![]()
图 1 胡杨3种异形叶特征
A:条形叶;B:卵形叶;C:锯齿阔卵形叶。
Figure 1. Characteristics of three heteromorphic leaves of Populus euphratica
A: Lanceolate leaf; B: Oval leaf; C: Serrated broad-oval leaf.
![]()
图 2 异形叶水势、相对含水量、水分饱和亏缺对地下水埋深梯度的响应
不同小写字母表示同一样地不同叶形间差异显著(P<0.05),不同大写字母表示同一叶形不同样地间差异显著(P<0.05);GWD:地下水埋深。下同。
Figure 2. Response of water potential, relative water content, and water saturation deficit in heteromorphic leaves to GWD
Different lowercase letters indicate significant differences among different leaf shapes in the same site (P<0.05), and different uppercase letters indicate significant differences among different sites in the same leaf (P<0.05); GWD: Groundwater depth. Same below.
![]()
图 3 异形叶渗透调节物质(Pro、SS、SP)含量对地下水埋深梯度的响应
Figure 3. Response of osmoregulatory substances (Pro, SS, SP) in heteromorphic leaves to GWD
![]()
图 4 异形叶MDA含量及SOD、POD活性对地下水埋深梯度的响应
Figure 4. Response of MDA content and SOD and POD activities in heteromorphic leaves to GWD
![]()
图 5 条形叶(A)、卵形叶(B)、锯齿阔卵形叶(C)生理指标的主成分分析
Figure 5. Principal component analysis of physiological traits of lanceolate (A), oval (B), and serrated broad-oval leaves (C)
表 1 样地概况
Table 1 Characteristics of study plots
样地
Site经纬度
Longitude and latitude地下水埋深
GWD / m密度
Density / ind./hm2平均树龄
Average age / a平均树高
Average height / m平均胸径
Average DBH / cm塔河源 40°30′04″N, 80°58′27″E 1.0~1.5 290.5 80.2 6.4 26.3 南口镇 40°26′29″N, 81°09′10″E 2.3~2.7 171.3 69.1 8.5 22.6 阿瓦提 40°19′44″N, 80°23′35″E 4.0~4.5 268.1 64.6 7.9 21.1 沙雅县 40°41′19″N, 81°59′43″E 7.5~8.0 120.2 110.6 7.8 36.4  下载: 导出CSV
下载: 导出CSV
表 2 条形叶生理指标与GWD的相关关系矩阵
Table 2 Correlation matrix of GWD and six physiological indicators of lanceolate leaves
指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.772** 0.709** −0.360 0.291 −0.194 −0.798** Pro 1.000 0.585* −0.379 0.733** 0.340 −0.754** SS 1.000 0.339 0.519 −0.299 −0.929** SP 1.000 0.119 −0.425 −0.159 MDA 1.000 0.440 −0.649* SOD 1.000 0.149 POD 1.000 注:*表示相关性显著(P<0.05),**表示相关性极显著(P<0.01)。下同。 Notes: *, significant (P<0.05); **, highly significant (P<0.01). Same below.
下载: 导出CSV
表 3 卵形叶生理指标与GWD的相关关系矩阵
Table 3 Correlation matrix of GWD and six physiological indicators of oval leaves
指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.441 0.617* 0.667* 0.098 −0.523 −0.799** Pro 1.000 0.342 0.801** 0.618* 0.395 −0.528 SS 1.000 0.760** 0.639* −0.628* −0.117 SP 1.000 0.720** −0.146 −0.389 MDA 1.000 0.082 0.174 SOD 1.000 0.034 POD 1.000
下载: 导出CSV
表 4 锯齿阔卵形叶生理指标与GWD的相关关系矩阵
Table 4 Correlation matrix of GWD and six physiological indicators of serrated broad-oval leaves
指标 Indicator GWD Pro SS SP MDA SOD POD GWD 1.000 0.392 0.840** 0.318 0.198 0.337 0.468 Pro 1.000 0.789** 0.188 0.707* 0.402 0.762** SS 1.000 0.122 0.621* 0.599* 0.784** SP 1.000 −0.430 −0.708* −0.216 MDA 1.000 0.760** 0.738** SOD 1.000 0.742** POD 1.000
下载: 导出CSV
-
[1] 代永欣,王林,万贤崇. 干旱导致树木死亡机制研究进展[J]. 生态学杂志,2015,34(11):3228−3236. Dai YX,Wang L,Wan XC. Progress on researches of drought-induced tree mortality mechanisms[J]. Chinese Journal of Ecology,2015,34(11):3228−3236.
[2] Anderegg WRL,Anderegg LDL. Hydraulic and carbohydrate changes in experimental drought-induced mortality of saplings in two conifer species[J]. Tree Physiol,2013,33(3):252−260. doi: 10.1093/treephys/tpt016
[3] Zhu GF,Li X,Su YH,Lu L,Huang CL. Seasonal fluctuations and temperature dependence in photosynthetic parameters and stomatal conductance at the leaf scale of Populus euphratica Oliv.[J]. Tree Physiol,2011,31(2):178−195. doi: 10.1093/treephys/tpr005
[4] 朱成刚,李卫红,马晓东,马建新,陈亚鹏. 塔里木河下游干旱胁迫下的胡杨叶绿素荧光特性研究[J]. 中国沙漠,2011,31(4):927−936. Zhu CG,Li WH,Ma XD,Ma JX,Chen YP. Chlorophyll fluorescence characteristic of Populus euphratica under drought stress in the lower reaches of Tarim River[J]. Journal of Desert Research,2011,31(4):927−936.
[5] Yordanov I,Velikova V,Tsonev T. Plant responses to drought,acclimation,and stress tolerance[J]. Photosynthetica,2000,38(2):171−186. doi: 10.1023/A:1007201411474
[6] Mittler R. ROS are good[J]. Trends Plant Sci,2017,22(1):11−19. doi: 10.1016/j.tplants.2016.08.002
[7] Luo WT,Zuo XA,Ma W,Xu C,Li A,et al. Differential responses of canopy nutrients to experimental drought along a natural aridity gradient[J]. Ecology,2018,99(10):2230−2239. doi: 10.1002/ecy.2444
[8] 岳喜元,左小安,庾强,徐翀,吕朋,张晶. 降水量和短期极端干旱对典型草原植物群落及优势种羊草(Leymus chinensis)叶性状的影响[J]. 中国沙漠,2018,38(5):1009−1016. Yue XY,Zuo XA,Yu Q,Xu C,Lü P,Zhang J. Effects of precipitation and short term extreme drought on leaf traits in inner Mongolia typical steppe[J]. Journal of Desert Research,2018,38(5):1009−1016.
[9] 陈图强,徐贵青,刘深思,米晓军,李彦. 干旱胁迫下新疆杨树冠不同高度叶片水分状况与非结构性碳动态[J]. 西北植物学报,2022,42(3):462−472. Chen TQ,Xu GQ,Liu SS,Mi XJ,Li Y. Leaf water status and non-structural carbohydrate dynamics with different crown height levels of Populus bolleana Lauche. under drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica,2022,42(3):462−472.
[10] 张旋,李蕊希,郑洲,李泽厚,贡璐,等. 极端干旱区多枝柽柳叶片功能性状及其与土壤理化因子的关系[J]. 生态学报,2023,43(9):3699−3708. Zhang X,Li RX,Zheng Z,Li ZH,Gong L,et al. Leaf functional traits of Tamarix ramosissima in extremely arid region and their relationship with soil physicochemical factors[J]. Acta Ecologica Sinica,2023,43(9):3699−3708.
[11] Wu X,Zheng XJ,Li Y,Xu GQ. Varying responses of two Haloxylon species to extreme drought and groundwater depth[J]. Environ Exp Bot,2019,158:63−72. doi: 10.1016/j.envexpbot.2018.11.014
[12] Bhusal N,Lee M,Han AR,Han A,Kim HS. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters[J]. Forest Ecol Manag,2020,465:118099. doi: 10.1016/j.foreco.2020.118099
[13] 郑清岭,杨忠仁,张凤兰,张晓艳,郝丽珍. 沙芥属植物活性氧清除系统对干旱胁迫的响应[J]. 西北植物学报,2018,38(9):1674−1682. Zheng QL,Yang ZR,Zhang FL,Zhang XY,Hao LZ. Responses of reactive oxygen species scavenging system to drought stress in Pugionium Gaertn.[J]. Acta Botanica Boreali-Occidentalia Sinica,2018,38(9):1674−1682.
[14] 郭艳阳,刘佳,朱亚利,柏延文,李红杰,等. 玉米叶片光合和抗氧化酶活性对干旱胁迫的响应[J]. 植物生理学报,2018,54(12):1839−1846. Guo YY,Liu J,Zhu YL,Bai YW,Li HJ,et al. Responses of photosynthetic and antioxidant enzyme activities in maize leaves to drought stress[J]. Plant Physiology Journal,2018,54(12):1839−1846.
[15] 王世绩. 全球胡杨林的现状及保护和恢复对策[J]. 世界林业研究,1996(6):37−44. Wang SJ. The status,conservation and recovery of global resources of Populus euphradica[J]. World Forestry Research,1996(6):37−44.
[16] Liu YB,Li XR,Chen GX,Li MM,Liu ML,Liu D. Epidermal micromorphology and mesophyll structure of Populus euphratica heteromorphic leaves at different development stages[J]. PLoS One,2015,10(9):e0137701. doi: 10.1371/journal.pone.0137701
[17] Yang XD,Anwar E,Xu YL,Zhou J,Sha LB,et al. Hydraulic constraints determine the distribution of heteromorphic leaves along plant vertical height[J]. Front Plant Sci,2022,13:941764. doi: 10.3389/fpls.2022.941764
[18] 苏培玺,张立新,杜明武,毕玉蓉,赵爱芬,刘新民. 胡杨不同叶形光合特性、水分利用效率及其对加富CO2的响应[J]. 植物生态学报,2003,27(1):34−40. doi: 10.17521/cjpe.2003.0005 Su PX,Zhang LX,Du MW,Bi YR,Zhao AF,Liu XM. Photosynthetic character and water use efficiency of different leaf shapes of Populus euphratica and their response to CO2 enrichment[J]. Chinese Journal of Plant Ecology,2003,27(1):34−40. doi: 10.17521/cjpe.2003.0005
[19] 祁鲁玉,陈浩楠,库丽洪·赛热别力,籍天宇,孟高德,等. 基于植物功能性状的暖温带5种灌木幼苗生长策略[J]. 植物生态学报,2022,46(11):1388−1399. doi: 10.17521/cjpe.2021.0396 Qi LY,Chen HN,Kulihong·Sairebieli,Ji TY,Meng GD,et al. Growth strategies of five shrub seedlings in warm temperate zone based on plant functional traits[J]. Chinese Journal of Plant Ecology,2022,46(11):1388−1399. doi: 10.17521/cjpe.2021.0396
[20] 黄文娟,李志军,杨赵平,白冠章. 胡杨异形叶结构型性状及其相互关系[J]. 生态学报,2010,30(17):4636−4642. Huang WJ,Li ZJ,Yang ZP,Bai GZ. The structural traits of Populus euphratica heteromorphic leaves and their correlations[J]. Acta Ecologica Sinica,2010,30(17):4636−4642.
[21] 杨扬,吕娜,郝建卿,李文海,白雪,郑彩霞. 胡杨枝条导水性与叶形变化关系研究[J]. 安徽农业科学,2010,38(20):11015−11017. Yang Y,Lü N,Hao JQ,Li WH,Bai X,Zheng CX. Preliminary research on relationship between hydraulic conduction property of branch and leaf shape change in Populus euphratica[J]. Journal of Anhui Agricultural Sciences,2010,38(20):11015−11017.
[22] 白雪,张淑静,郑彩霞,郝建卿,李文海,杨扬. 胡杨多态叶光合和水分生理的比较[J]. 北京林业大学学报,2011,33(6):47−52. Bai X,Zhang SJ,Zheng CX,Hao JQ,Li HW,Yang Y. Comparative study on photosynthesis and water physiology of polymorphic leaves of Populus euphratica[J]. Journal of Beijing Forestry University,2011,33(6):47−52.
[23] 王海珍,韩路,徐雅丽,刘艳萍,王琳. 胡杨异形叶光合系统Ⅱ叶绿素荧光特性[J]. 西北植物学报,2019,39(10):1795−1804. Wang HZ,Han L,Xu YL,Liu YP,Wang L. Chlorophyll fluorescence characteristics of photosystem Ⅱ of Populus euphratica heteromorphic leaves[J]. Acta Botanica Boreali-Occidentalia Sinica,2019,39(10):1795−1804.
[24] 尹秋龙. 黄土高原木本植物叶经济性状和水力性状研究[D]. 西安: 西北大学, 2019: 15-32. [25] 秦少伟,李才林,吴海柱,闫伟伟,何志贵,等. 次生代谢相关ceRNA调控网络在胡杨异形叶发生中的作用分析[J]. 分子植物育种,2023,21(10):3253−3263. Qin SW,Li CL,Wu HZ,Yan WW,He ZG,et al. The role of ceRNA regulatory networks related to secondary metabolism in the morphogenesis of Populus euphratica heterophyll[J]. Molecular Plant Breeding,2023,21(10):3253−3263.
[26] 韩路,王海珍,牛建龙,王家强,柳维扬. 荒漠河岸林胡杨群落特征对地下水位梯度的响应[J]. 生态学报,2017,37(20):6836−6846. Han L,Wang HZ,Niu JL,Wang JQ,Liu WY. Response of Populus euphratica communities in a desert riparian forest to the groundwater level gradient in the Tarim Basin[J]. Acta Ecologica Sinica,2017,37(20):6836−6846.
[27] 柏新富,卜庆梅,谭永芹,朱建军. 植物4种水势测定方法的比较及可靠性分析[J]. 林业科学,2012,48(12):128−133. Bai XF,Bu QM,Tan YQ,Zhu JJ. Comparison and analysis of four methods used in measuring the plant water potential[J]. Scientia Silvae Sinicae,2012,48(12):128−133.
[28] 姜华,毕玉芬,陈连仙,单贵莲,何承刚. 旱作条件下紫花苜蓿生理特性的研究[J]. 草地学报,2012,20(6):1077−1080. Jiang H,Bi YF,Chen LX,Shan GL,He CG. Physiological characteristics of alfalfa under dry-farming conditions[J]. Acta Agrestia Sinica,2012,20(6):1077−1080.
[29] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006: 210-217. [30] 孙启兴,杨晓东,李浡睿,孔翠翠,伊力哈穆江·艾尼弯,等. 水力性状对荒漠植物群落物种多度分布格局的影响[J]. 干旱区研究,2023,40(3):412−424. Sun QX,Yang XD,Li BR,Kong CC,Elhamjan·Anwar,et al. Effects of hydraulic traits on the species abundance distribution pattern of desert plant communities[J]. Arid Zone Research,2023,40(3):412−424.
[31] Villagra M,Campanello PI,Bucci SJ,Goldstein G. Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species[J]. Tree Physiol,2013,33(12):1308−1318. doi: 10.1093/treephys/tpt098
[32] Meixner M,Foerst P,Windt CW. Reduced spatial resolution MRI suffices to image and quantify drought induced embolism formation in trees[J]. Plant Methods,2021,17(1):38. doi: 10.1186/s13007-021-00732-7
[33] 高亚宁, 常娇, 李玉, 姜元昊, 轩正英, 阿依买木·沙吾提. 干旱胁迫对芜菁苗期生理生化的影响及抗旱性评价[J/OL]. 分子植物育种, 2023. http://kns.cnki.net/kcms/detail/46.1068.S.20230303.1347.009.html. Gao YN, Chang J, Li Y, Jiang YH, Xuan ZY, Ayimaimu · Shawuti. Effect of drought on physiological and biochemistry of turnip seedlings and evaluation of drought resistance[J/OL]. Molecular Plant Breeding, 2023. http://kns.cnki.net/kcms/detail/46.1068.S.20230303.1347.009.html.
[34] 童德龙,徐雅丽,焦培培. 土壤水分胁迫对胡杨、灰叶胡杨渗透调节物质和保护酶活性的影响[J]. 江苏农业科学,2013,41(1):169−173. [35] 马步东,黄海霞,陆刚,周晓瑾,张君霞. 土壤干旱过程对裸果木荧光特性及渗透调节物质的影响[J]. 西北林学院学报,2022,37(3):30−36. Ma BD,Huang HX,Lu G,Zhou XJ,Zhang JX. Effects of soil drought on fluorescence characteristics and osmotic regulatory substances of Gymnocarpos przewalskii[J]. Journal of Northwest Forestry University,2022,37(3):30−36.
[36] 韩蕊莲,李丽霞,梁宗锁. 干旱胁迫下沙棘叶片细胞膜透性与渗透调节物质研究[J]. 西北植物学报,2003,23(1):23−27. Han RL,Li LX,Liang ZS. Seabuckthorn relative membrane conductivity and osmotic adjustment under drought stress[J]. Acta Botanica Boreali-Occidentalia Sinica,2003,23(1):23−27.
[37] Sharma SS,Dietz KJ. The relationship between metal toxicity and cellular redox imbalance[J]. Trends Plant Sci,2009,14(1):43−50. doi: 10.1016/j.tplants.2008.10.007
[38] 吕朝燕,高智席,徐兴线,张谣,陈旭,石娇娇. 干旱胁迫及复水对桫椤生理特性的影响[J]. 林业资源管理,2022(5):160−168. Lü CY,Gao ZX,Xu XX,Zhang Y,Chen X,Shi JJ. Effects of drought stress and rehydration on the physiological characteristics of Alsophila spinulosa[J]. Forest Resources Management,2022(5):160−168.
[39] 李萍萍,曾明,李文海,赵媛媛,郑彩霞. 胡杨异形叶抗氧化能力的比较[J]. 北京林业大学学报,2019,41(8):76−83. Li PP,Zeng M,Li WH,Zhao YY,Zheng CX. Comparative study on antioxidant capacity of heteromorphic leaves of Populus euphratica[J]. Journal of Beijing Forestry University,2019,41(8):76−83.
[40] Pour-Aboughadareh A,Omidi M,Naghavi MR,Etminan A,Mehrabi AA,Poczai P. Wild relatives of wheat respond well to water deficit stress:a comparative study of antioxidant enzyme activities and their encoding gene expression[J]. Agriculture,2020,10(9):415. doi: 10.3390/agriculture10090415
[41] 王孟本,冯彩平,李洪建,柴宝峰,武冬梅. 树种保护酶活性与PV曲线水分参数变化的关系[J]. 生态学报,2000,20(1):173−176. Wang MB,Feng CP,Li HJ,Chai BF,Wu DM. The relationship between the activities of protecting enzymes and water parameters from pressure-volume curves in six tree species[J]. Acta Ecologica Sinica,2000,20(1):173−176.
[42] 李端,司建华,李继彦,王佩将,原黎明. 胡杨(Populus euphratica)对盐胁迫和干旱胁迫的生理响应特征[J]. 中国沙漠,2023,43(2):205−215. Li D,Si JH,Li JY,Wang PJ,Yuan LM. Physiological responses and differences of Populus euphratica to salt stress and drought stress[J]. Journal of Desert Research,2023,43(2):205−215.
[43] 王海珍,韩路,徐雅丽,王琳. 控水处理对胡杨、灰胡杨生长特性及水分利用效率的影响[J]. 农业工程学报,2010,26(S2):195−201. Wang HZ,Han L,Xu YL,Wang L. Effects of water stress on growth characteristics and water use efficiency of Populus euphratica and Populus pruinosa[J]. Transactions of the Chinese Society of Agricultural Engineering,2010,26(S2):195−201.
-
期刊类型引用(1)
1. 何拓,曾岩,殷亚方,张坤,袁良琛,董晖,周志华. 为野生植物保护和可持续贸易奠定科学基础——CITES植物委员会第27次会议评述. 生物多样性. 2024(09): 171-175 .  百度学术
百度学术
其他类型引用(0)


计量
- 文章访问数: 1160
- HTML全文浏览量: 1272
- PDF下载量: 16
- 被引次数: 1
