
Floristic characteristics of seed plant species in valley forests of the Irtysh River basin in Xinjiang
-
摘要:
基于第三次新疆综合科学考察的实地调查数据,整理了额尔齐斯河流域河谷林种子植物名录,并对河谷林种子植物物种多样性及区系特征进行了系统分析。结果显示,额尔齐斯河流域河谷林共记录种子植物55科228属403种,分别占新疆种子植物科、属、种总数的48.67%、29.73%、10.84%,植物多样性相对丰富。河谷林草本植物种类丰富,占种子植物总数的80.40%,乔木物种较单一,其中杨柳科是群落优势建群科和特征科,灌木植物缺乏是该区重要特点。河谷林种子植物区系组成及来源复杂,科和属的分布区类型以世界广布最多,分别为科、属总数的60.00%和19.74%,科和属的温带性成分占非世界广布类型的59.09%和92.90%,以温带成分占明显优势。河谷林具有明显的过渡特征,反映在物种层面既有适应干旱环境的旱生或超旱生植物,又有适应山地森林、草甸和湿地的植物的过渡性特征,因此,河谷林是区域重要的植物种库,保护价值高。依据群落物种组成、优势树种和地形地貌差异,建群种组成差异及干扰程度,可将河谷林划分为山前河谷林与山地河谷林两个类型。
Abstract:Based on a field survey conducted during the Third Xinjiang Scientific Expedition, this study compiled a comprehensive list of seed plants in the valley forests of the Irtysh River basin in Xinjiang and analyzed their floristic characteristics using floristic geography methods. Results identified 403 seed plants belonging to 55 families and 228 genera, representing 48.67%, 29.73%, and 10.84% of seed plant families, genera, and species in Xinjiang, respectively, highlighting rich plant diversity. Herbaceous plants dominated the flora, accounting for 80.40% of the total species, whereas tree species were relatively limited, with Salicaceae serving as the dominant constructive and characteristic family within the communities. Furthermore, shrubs were notably absent, representing an important aspect of the valley forests. Geographically, the families and genera were classified into seven and 25 distribution types, respectively, with 59.09% of non-cosmopolitan families and 92.90% of non-cosmopolitan genera exhibiting temperate distribution patterns. The interaction between plant species and environmental factors has resulted in transitional adaptations, including xerophytes and hyper-xerophytes suited to arid conditions and cold-adapted hygrophytes thriving in mountain forests and alpine meadows. These valley forests act as a crucial regional species pool with significant conservation value. Moreover, the study also revealed that valley forests can be classified as piedmont or mountain according to community composition, dominant tree species, topography, and geomorphology.
-
Keywords:
- Irtysh River basin /
- Valley forests /
- Seed plant /
- Flora /
- Species diversity
-
高寒草地是青藏高原最主要的生态系统,是维护国家生态安全和改善青藏高原地区农牧民生活质量的重要保障[1]。在过去几十年里,受人类活动和自然因素的影响,高寒草地的生产和生态功能急剧下降,造成大面积土地裸露和草畜不平衡等诸多问题[2]。最近,经过相关领域学者们的不断探索实践,我国高寒草地退化的趋势明显改善,植被覆盖度大幅提升,水土流失和荒漠化基本遏制[3]。然而,青藏高原地区气候寒冷,有机质分解较慢,土壤微生物活性较低[4],导致退化草地治理周期较长,草畜矛盾仍未能有效解决。因此,在退化草地恢复的同时,建植多年生人工草地便成为满足生产和生态功能需求的重要手段。在实践中发现,多年生高寒栽培草地在建植2~3年后地上生物量达到峰值,第4年开始其地上生物量逐年下降,且饲草品质普遍较低[5]。究其原因,与青藏高原地区土壤氮限制密切相关[6]。青藏高原地区土壤氮限制极其严重,加之气温较低,严重抑制了与氮有关的功能微生物的活性,导致生产力较低[6]。此外,多年生牧草常年吸收土壤中的营养物质,导致土壤养分贫瘠,进而造成草地减产[7]。综合两方面的原因,氮添加成为提升多年生栽培草地生产功能的直接手段。
氮素形态和施氮水平是草地氮肥管理的两项重要内容,对植物生长发育起着关键调节作用。根据化合物形态可将氮肥分为铵态氮肥(A)、酰胺态氮肥(U)和硝态氮肥(N)等多种类型,不同形态氮肥均能促进植物的生长,但在土壤中的转化机制不同,导致植物的干物质分配和营养积累存在差异[8]。植物可以吸收利用的土壤氮素主要为硝态氮和铵态氮,氮素形态对不同植物生长发育的影响存在差异,氮肥形态是影响肥效的主要因素之一[9]。施用不同形态氮素会影响作物的根系发育及碳氮代谢等生理进程,进而影响植株生物量的积累。同时,氮素形态还会影响糖、激素、维生素和生物碱等各种化学物质的合成,进而影响植物的品质[10]。向雪梅等[11]在高寒区的施氮研究表明,酰胺态氮能保证植物较高的氮素利用率和较低的氮损失率,是提高植物生产力最佳的氮素形态,而Guo等[12]研究发现,硝态氮能促进野牛草(Buchloe dactyloides (Nutt.) Engelm.)的营养繁殖和品质积累。两种结果不一致的主要原因是不同植物对氮素形态的需求存在差异,故应根据植物类型和生存环境设置合理的氮源。对于青藏高原地区多年生高寒栽培草地而言,哪种氮素形态更有利于生产力和营养品质的积累不得而知。此外,研究施氮水平对植物生产性能的影响并确定合理的氮添加量也是科学添加氮肥的关键[13]。以往研究证实,合理的氮添加量是保证植物生产力和营养品质的关键[14],但关于具体氮添加量始终没有形成统一的结论,这是因为施氮水平应符合环境状况,不同区域氮素水平的阈值存在显著差异,氮限制比较严重的土壤环境可能需要施加更多的氮肥来满足植物的生长需求[15]。但过量的氮添加不仅造成氮肥利用率下降,也会对生态环境构成威胁。因此,确定最佳施氮量也是维持多年生高寒栽培草地生产力和营养品质的核心目标。
鉴于此,本研究以4年龄人工草地为研究对象,设置3个氮素形态和4个施氮水平,通过比较不同处理植物的地上生物量和饲草营养品质等参数,探究不同氮素形态和施氮水平对饲草生产性能和营养品质的影响,通过灰色关联度综合分析,进一步筛选出最佳的氮素形态和施氮水平,以期为高寒地区优质饲草的生产提供科学依据。
1. 材料与方法
1.1 试验区概况
试验区位于青海省海南藏族自治州共和县巴卡台农牧场(36°17'N,100°55'E),平均海拔3 300 m,年均降水量为300 mm,年蒸发量在2 000~2 400 mm,年平均气温为4.1 ℃。试验区冬季寒冷漫长,夏季温和短暂,年内干旱少雨且温度偏低,气温垂直分布明显,太阳辐射强,属高原大陆性气候特征。降水季节性分布不均,主要集中在7-10月。试验地土壤为高山草甸土和黄绵土[16]。
1.2 试验设计
于2022年6月,以2019年建植的青海草地早熟禾(Poa pratensis L. cv. Qinghai)+青海中华羊茅(Festuca sinensis L. cv. Qinghai)混播草地为研究对象,两个物种的混播比例为1∶1,建植面积为15 m×150 m。设置小区时,为避免边际效应,选取整块地中间较均匀的地段,采用随机区组设计,设置3个氮素形态,分别为尿素(酰胺态氮,U)、硫酸铵(铵态氮,A)和硝酸钙(硝态氮,N);参照中国氮沉降分布格局(青海地区干湿沉降率7.55 kg·hm−2·yr−1)确定氮素添加剂量,设4个施氮梯度,分别为青海省干湿氮沉降的0、3、6、9倍,浓度依次为0(T0,CK)、22.5(T1)、45(T2)、67.5 kg·hm−2·a−1(T3),各处理见表1。每个处理3个重复,共30个小区,小区面积为4 m×4 m,小区之间间隔5 m。将称好的肥料分为两等份,分别于6月上旬和下旬加到2 L水中溶解,摇匀后装入喷壶,均匀喷洒在相应的小区内,CK处理喷洒相同体积的水。
表 1 氮素形态和施氮水平设置Table 1. Nitrogen forms and nitrogen level settings施肥处理
Fertilization treatment氮素形态
Nitrogen form施氮水平
Nitrogen level / kg·hm−2·a−1T0(CK) − 0.0 UT1 酰胺态氮 22.5 UT2 45.0 UT3 67.5 AT1 铵态氮 22.5 AT2 45.0 AT3 67.5 NT1 硝态氮 22.5 NT2 45.0 NT3 67.5 1.3 样品采集与分析
1.3.1 样品采集与处理
于2022年8月上旬(植物生长旺期)对各试验小区进行调查与样品采集。采用样方法,设置50 cm×50 cm的样方,齐地面刈割后带回实验室称取鲜重,后转移至105 ℃烘箱中杀青30 min,然后在75 ℃烘干至恒重,称取地上生物量。之后将烘干草样用粉碎机粉碎,过1 mm筛备用。
1.3.2 牧草品质测定方法
利用元素分析仪测定植物粗蛋白含量[17],利用索氏抽提法测定粗脂肪含量[18],使用马弗炉灼烧法测定粗灰分含量[18],中性洗涤纤维含量和酸性洗涤纤维含量采用范式纤维洗涤法进行测定[19],并根据童永尚等[19]的方法计算相对饲喂价值。
1.3.3 数据处理与分析
采用Excel 2010软件进行原始数据整理和灰色关联度分析,具体参见童永尚等[19]的方法。运用SPSS 27.0软件进行双因素方差分析和聚类分析。在Origin 2022软件中绘图。
2. 结果与分析
2.1 氮素形态和施氮水平对饲草生产性能的影响
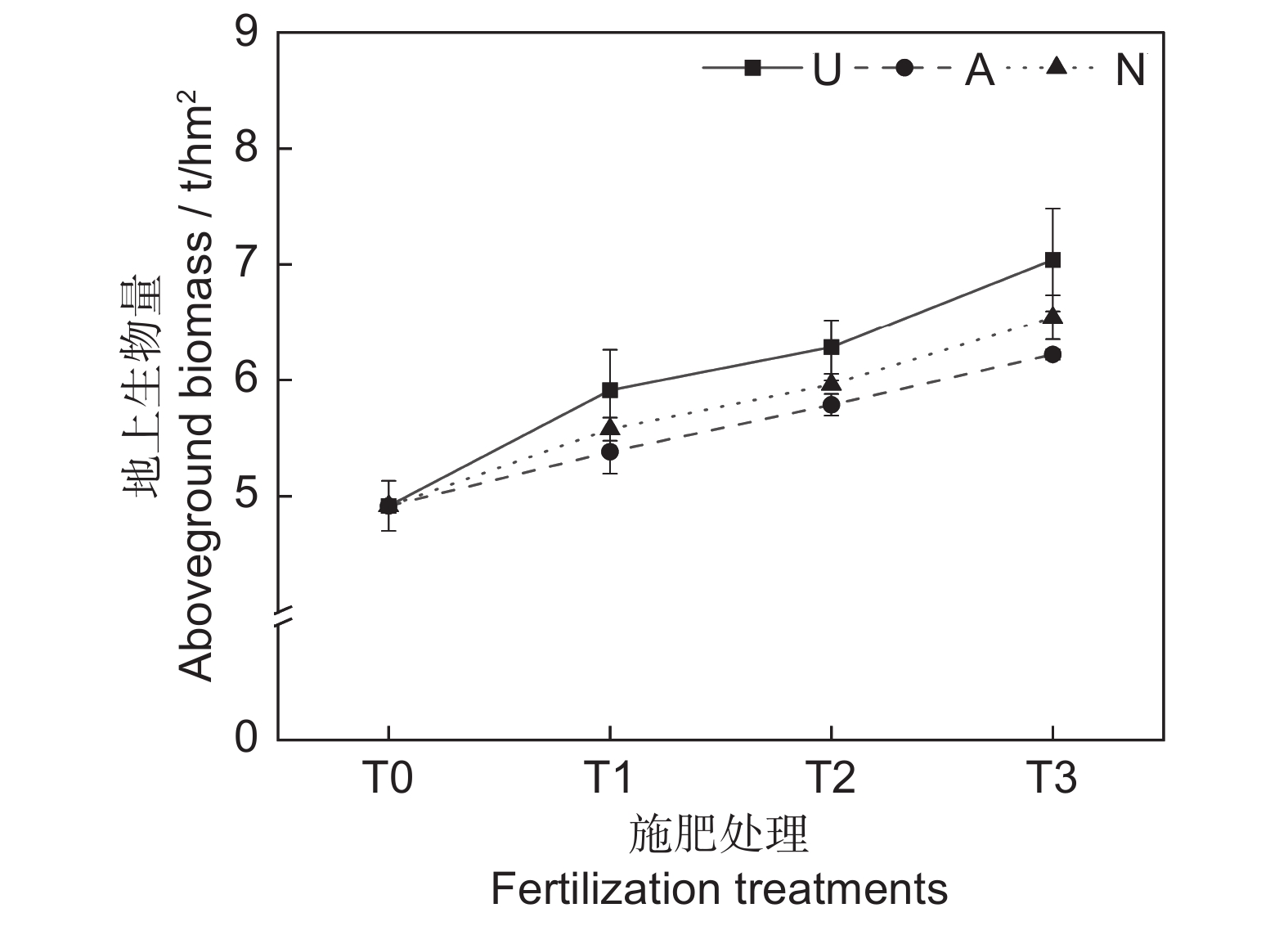
由表2可知,氮素形态对多年生高寒栽培草地的地上生物量影响显著(P<0.05),施氮水平对地上生物量的影响极显著(P<0.001),氮素形态和施氮水平交互作用对地上生物量影响不显著。氮添加整体上提高了群落地上生物量,且随着施氮水平的提高呈递增趋势,UT3、AT3和NT3处理分别较CK处理提高43.22%、26.54%和33.11%。同等氮水平相比较,对地上生物量的促进作用表现为U>N>A(图1)。
表 2 氮素形态和施氮水平的双因素方差分析Table 2. Two factor analysis of variance for nitrogen forms and nitrogen application levels影响因素
Influence factor氮素形态
Nitrogen form (F)施氮水平
Nitrogen level (L)氮素形态×施氮水平
F×LF P F P F P 地上生物量 5.622 <0.05 14.056 <0.001 0.153 0.959 粗蛋白 18.073 <0.001 38.120 <0.001 1.748 0.184 粗脂肪 39.829 <0.001 5.377 <0.05 1.183 0.351 粗灰分 0.020 0.980 0.576 0.572 1.714 0.191 中性洗涤纤维 1.979 0.167 1.423 0.267 1.422 0.267 酸性洗涤纤维 1.153 0.338 0.718 0.501 1.555 0.229 相对饲喂价值 1.275 0.304 1.284 0.301 1.909 0.153 2.2 氮素形态和施氮水平对饲草营养品质的影响
2.2.1 对饲草粗蛋白含量的影响
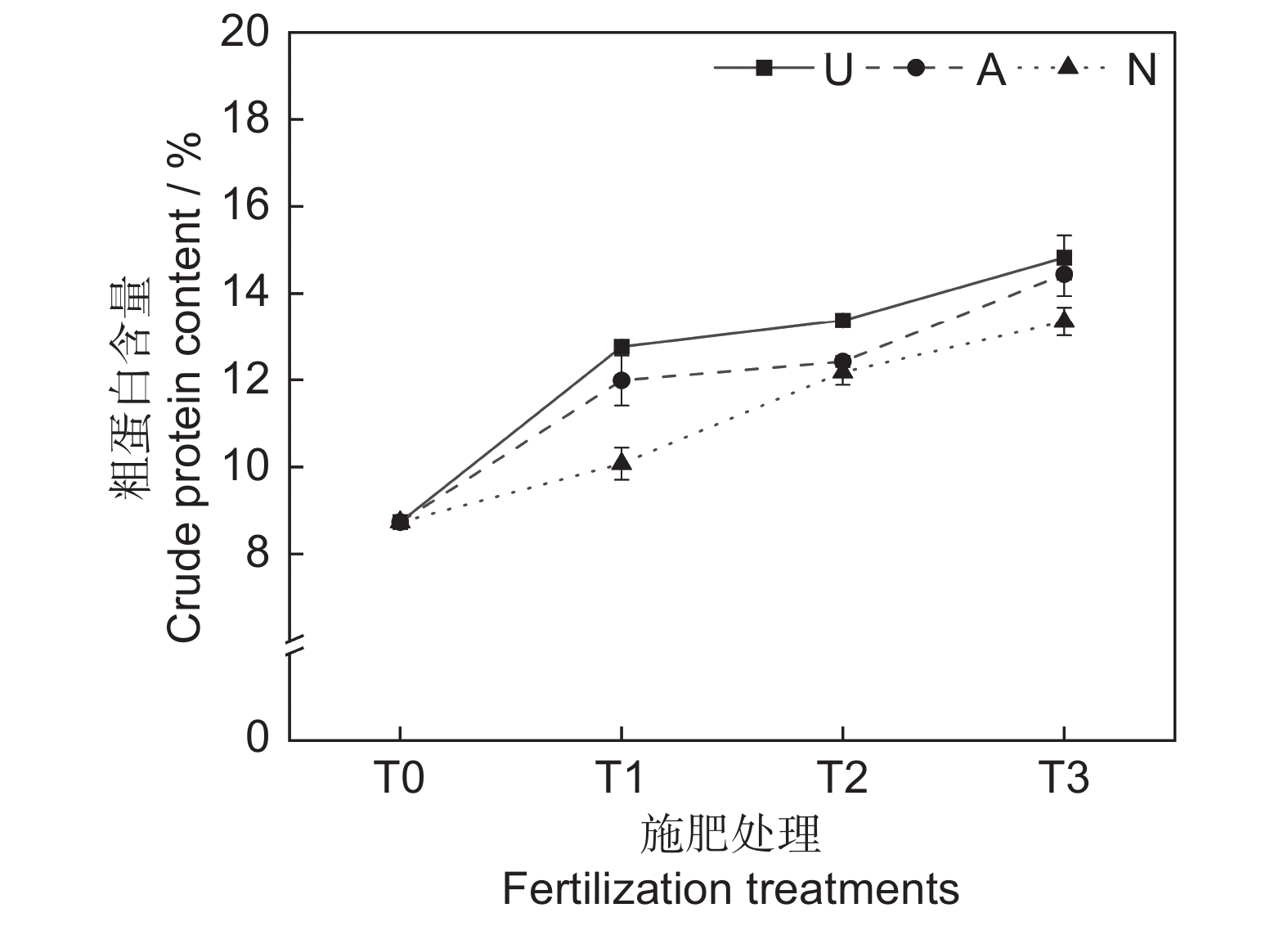
由表2和图2可知,氮素形态和施氮水平对饲草粗蛋白含量均具有极显著影响(P<0.001),氮素形态和施氮水平交互作用对饲草粗蛋白含量影响不显著。氮添加整体上提高了植物的粗蛋白含量,且随着施氮水平的提高呈递增趋势,UT3、AT3和NT3处理分别较CK处理提高了69.76%、65.41%和52.86%。同等氮水平相比较,对饲草粗蛋白含量的促进作用表现为U>A>N。
2.2.2 对饲草粗脂肪含量的影响
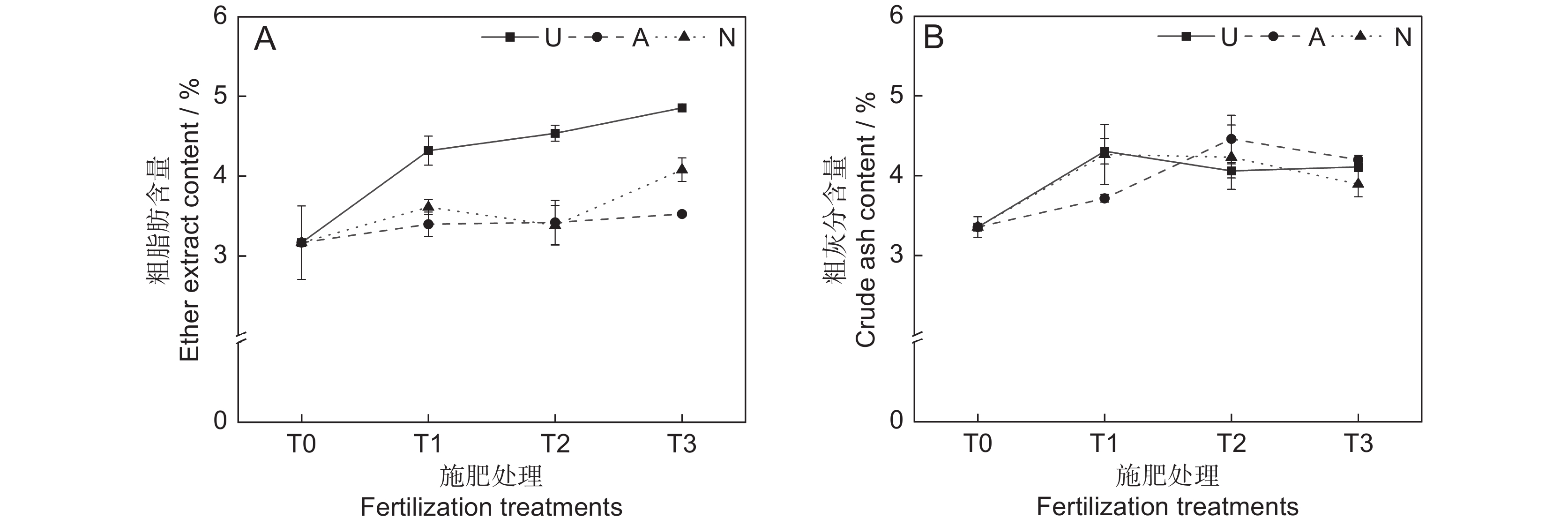
氮素形态对饲草粗脂肪含量具有极显著影响(P<0.001),施氮水平对饲草粗脂肪含量具有显著影响(P<0.05),氮素形态和施氮水平对饲草粗脂肪含量没有明显的交互作用(表2)。氮添加整体上提高了植物的粗脂肪含量,不同施氮水平对饲草粗脂肪含量的促进作用因氮素形态不同而有所差异。饲草粗脂肪含量随着酰胺态氮施肥水平的提高而增加,铵态氮施肥水平的增加对植物粗脂肪含量的影响较小。3种氮素形态下,饲草的粗脂肪含量均在T3水平时达到峰值,UT3、AT3和NT3处理分别较CK提高了53.30%、11.34%和28.86%。同等氮水平相比较,对地上生物量的促进作用表现为U>N>A(图3:A)。
![]() 图 3 不同处理下饲草粗脂肪(A)和粗灰分(B)含量Figure 3. Ether extract (A) and crude ash (B) content of forage under different treatments
图 3 不同处理下饲草粗脂肪(A)和粗灰分(B)含量Figure 3. Ether extract (A) and crude ash (B) content of forage under different treatments2.2.3 对饲草粗灰分含量的影响
由表2可知,氮素形态、施氮水平及二者交互作用对饲草粗灰分含量的影响均不显著。氮添加整体上提高了植物的粗灰分含量(P<0.05),AT2处理下植物的粗灰分含量最高,为4.46%。同一氮素形态下不同施氮水平相比较,A和N处理均在T1水平时达到最大粗灰分含量,分别为4.31%和4.26%(图3:B)。
2.2.4 对饲草中性洗涤纤维含量的影响
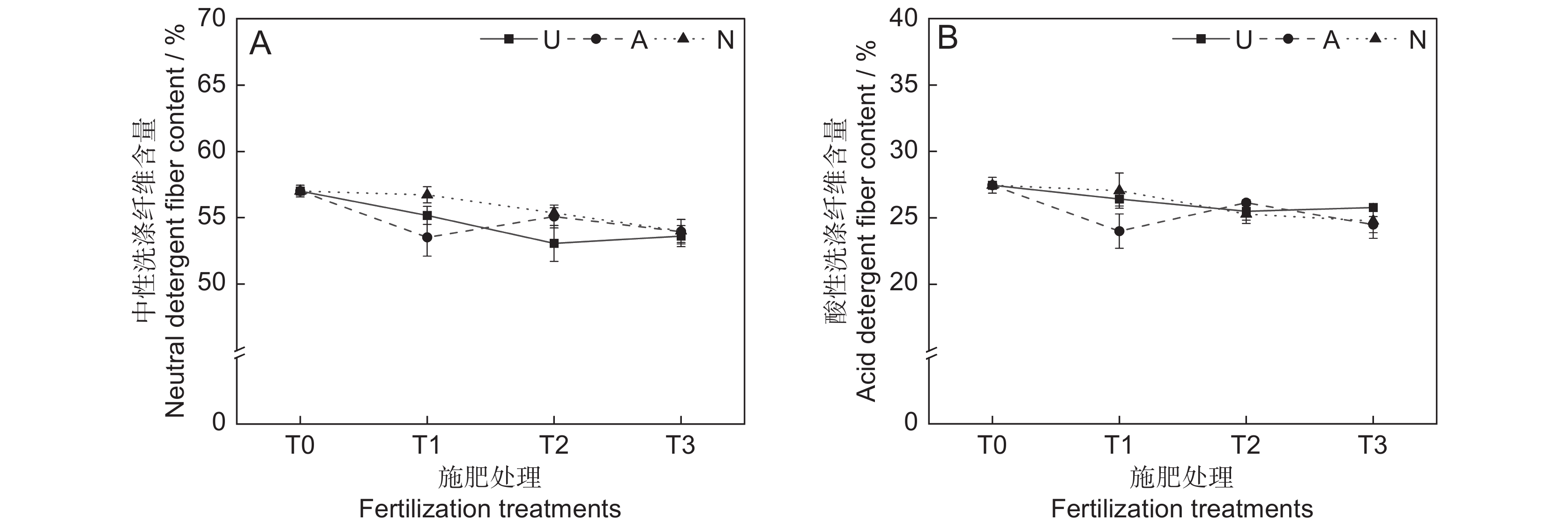
本研究发现,氮素形态、施氮水平及二者交互作用对饲草中性洗涤纤维含量均未产生显著影响(表2)。氮添加整体上降低了植物的中性洗涤纤维含量(P<0.05)。随着硝态氮施肥水平的提高,植物中性洗涤纤维含量逐渐降低(图4:A)。3种氮肥在T3水平时的中性洗涤纤维含量趋于一致,介于63.6%~64.0%。
![]() 图 4 不同处理下饲草中性洗涤纤维(A)和酸性纤维(B)含量Figure 4. Neutral detergent fiber (A) and acid detergent fiber (B) content of forage under different treatments
图 4 不同处理下饲草中性洗涤纤维(A)和酸性纤维(B)含量Figure 4. Neutral detergent fiber (A) and acid detergent fiber (B) content of forage under different treatments2.2.5 对饲草酸性洗涤纤维含量的影响
由表2可知,氮素形态、施氮水平、氮素形态和施氮水平交互作用对饲草酸性洗涤纤维含量的影响均不显著。氮添加整体上降低了植物的酸性洗涤纤维含量(图4:B)。T2水平时,3种氮肥对植物酸性洗涤纤维含量的影响不明显。总体来看,T1水平时,施用铵态氮肥对植物酸性洗涤纤维含量的降低效果最为明显。
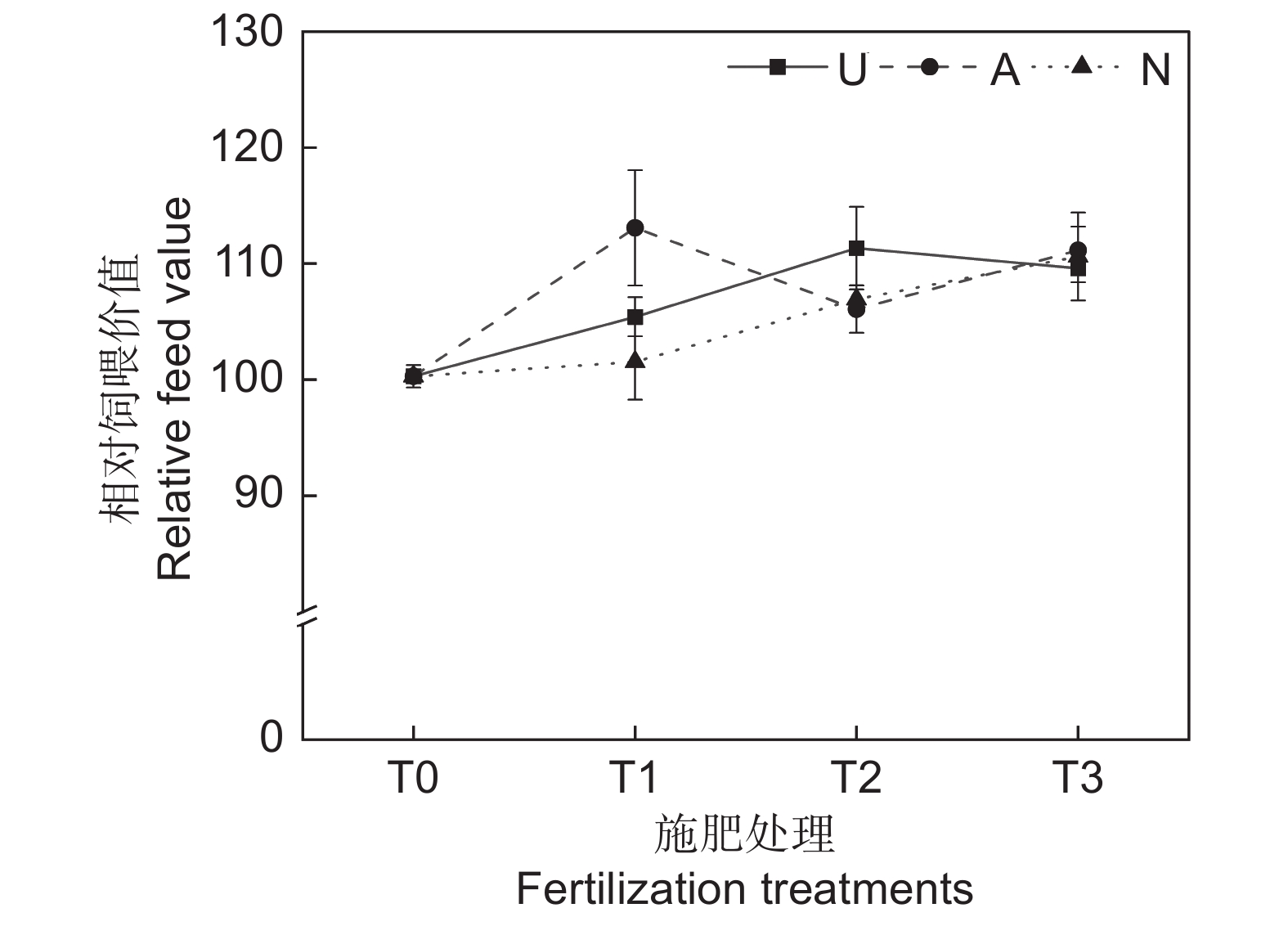
2.2.6 对饲草相对饲喂价值的影响
由表2可知,氮素形态、施氮水平及二者交互作用对饲草相对饲喂价值均未产生显著影响。如图5所示,氮添加整体上提高了饲草的相对饲喂价值,不同氮素形态和施氮水平对饲草相对饲喂价值的影响较大。随着硝态氮肥施氮水平的提高,饲草的相对饲喂价值逐渐提高。T3水平时,3种氮肥对饲草相对饲喂价值的提升效果接近。不同氮肥形态相比较,T1水平时,施用铵态氮肥的饲草其相对饲喂价值最高,为113.08;T2水平时,施用酰胺态氮肥时饲草的相对饲喂价值最高,为113.34。
![]() 图 5 不同处理下饲草相对饲喂价值Figure 5. Relative feeding value of forage under different treatments
图 5 不同处理下饲草相对饲喂价值Figure 5. Relative feeding value of forage under different treatments2.3 灰色关联度评价和聚类分析
将植物地上生物量、粗蛋白含量和中性洗涤纤维含量等7个指标纳入评价体系,对不同氮素形态和施肥水平共10个处理进行灰色关联度综合评价。由表3可知,CK处理排序第9,说明氮添加普遍提高了饲草的综合性能。综合排名前4位的处理分别是UT3、UT2、NT3和AT3。不同酰胺态氮肥处理的综合排名依次为UT3>UT2>UT1,不同铵态氮肥处理的综合排名依次为AT3>AT1>AT2,不同硝态氮肥处理的综合排名依次为NT3>NT2>NT1。
表 3 不同处理的灰色关联度综合评价Table 3. Comprehensive evaluation of gray correlation degree for different treatments施肥处理
Fertilization treatment等权关联度
Gray correlative排序
Rank权重系数
Weight coefficient加权关联度
Weighted gray correlative加权关联度排序
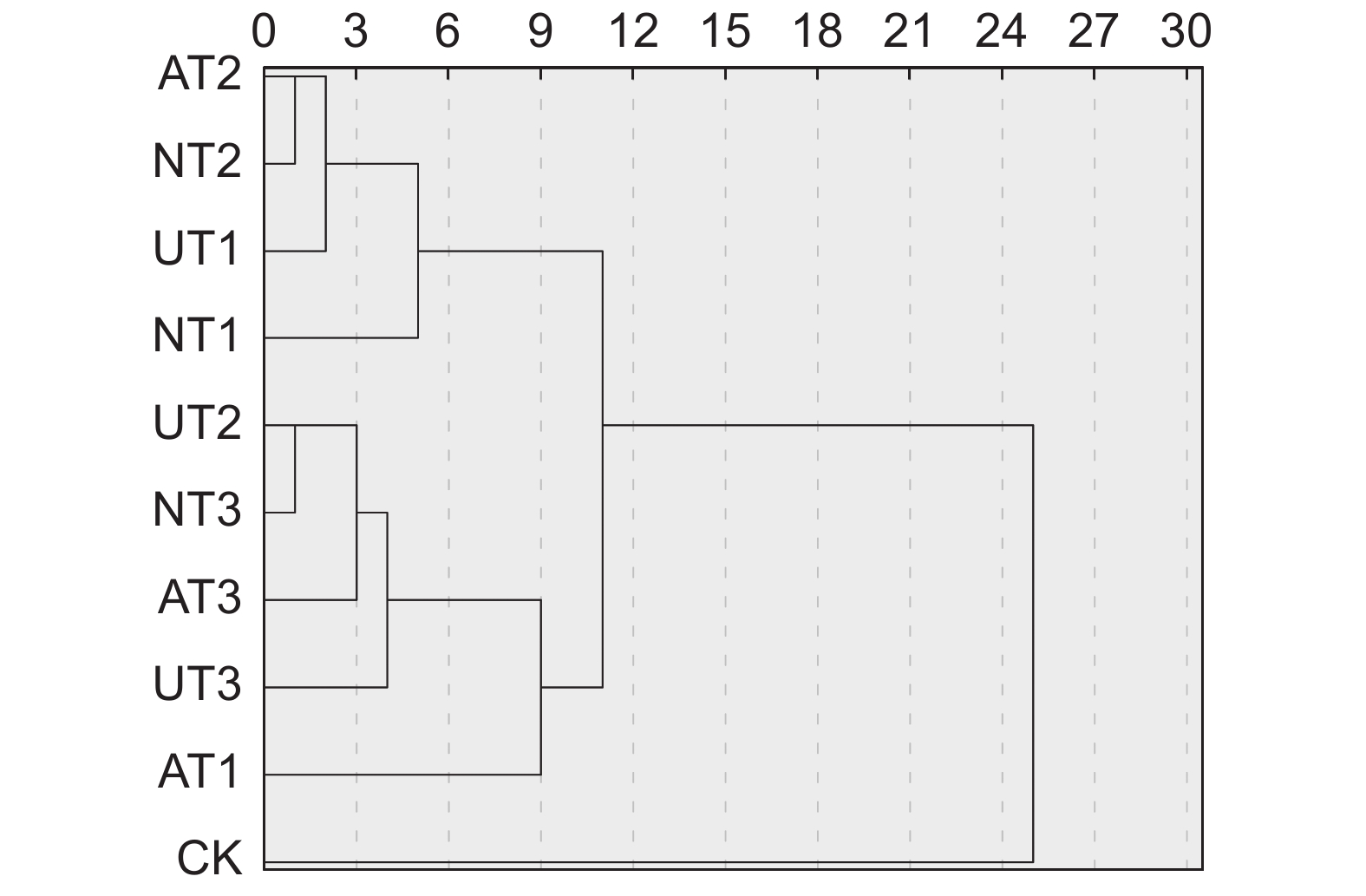
Rank of weighted gray correlativeUT3 0.871 4 1 0.126 3 0.110 0 1 UT2 0.763 3 2 0.110 6 0.084 4 2 NT3 0.753 6 3 0.109 2 0.082 3 3 AT3 0.746 1 4 0.108 1 0.080 7 4 AT1 0.718 6 5 0.104 1 0.074 8 5 UT1 0.654 1 6 0.094 8 0.062 0 6 NT2 0.633 7 7 0.091 8 0.058 2 7 AT2 0.613 0 8 0.088 8 0.054 5 8 CK 0.589 1 9 0.085 4 0.050 3 9 NT1 0.557 0 10 0.080 7 0.045 0 10 本试验将10个处理的地上生物量和营养品质进行了聚类分析,运用SPSS 27.0软件构建树形图(图6),在欧式距离为9处,可将其分为4大类。第Ⅰ类仅包括CK处理,其产量和营养品质均表现最差;第Ⅱ类只有AT1处理,其地上生物量略高于CK处理;第Ⅲ类包括UT2、UT3、AT3和NT3处理,其产量和营养品质均表现最好;第Ⅳ类包括UT1、NT1、NT2和AT2处理,其产量和营养品质仅次于第Ⅲ类。
3. 讨论
3.1 氮素形态和施氮水平对饲草生产性能的影响
众所周知,氮添加可以提高植物的生产力,但不同环境的氮添加量应有所差异[20]。施氮量较低时,土壤养分仍然不能满足植物生长,施氮量过多又会导致土壤中产生有毒的亚硝酸盐[21],进而阻碍植物生长,因此栽培草地施氮量应根据土壤氮素含量来确定。在以往的研究中,关于氮添加水平对植物地上生物量的调节作用有两种结论,第1种是地上生物量随施氮量增加而增加[22];第2种是地上生物量随施氮量的增加会出现一个峰值,然后逐渐降低[15],这是因为氮添加量有一定的环境阈值,存在一个最适施氮量,以最适施氮量为对称轴,植物的地上生物量随施氮量增加呈抛物线变化。本研究中,施氮水平对地上生物量具有极显著影响,氮添加显著提高了群落地上生物量,且随着施氮水平的提高呈递增趋势,在施氮量为67.5 kg·hm−2·a−1时达到峰值,说明本研究中设置的施氮量可能过低,在后期的试验中需要加大施氮梯度。此外,氮素形态对植物的地上生物量也产生显著影响,对地上生物量的促进作用表现为U>N>A。说明在短期内,酰胺态氮对植物生物量的促进效果更为明显,而硝态氮和铵态氮的效果相对较差,这与向雪梅等[11]和芦光新等[23]在高寒草地中的研究结论相似。尽管施加铵态氮肥补充了土壤养分,一定程度上促进了植物生长,但铵态氮肥抑制了植物对K+和Ca2+的吸收,导致NH4+的积累并产生氨害,从而限制了植物的生长[12, 24]。硝态氮肥的促产作用介于酰胺态和铵态氮肥之间。硝态氮同样也会对植物的生长产生不利影响,在还原过程中,硝态氮会消耗较多的能量。另外,在弱光条件下,植物对硝态氮的吸收也有可能会受到抑制,从而导致氮素供应不足。相比铵态氮肥和硝态氮肥,含氮量较高的酰胺态氮肥为土壤补充了充足的养分[16],更能满足植物的生长需求。由于氮素形态和氮素水平的设置还与牧草栽培方式、施氮时间和气候条件等因素相关,后期还应针对以上干扰因素设置控制试验,进行深入研究。
3.2 氮素形态和施氮水平对饲草营养品质的影响
在高寒地区,老龄人工草地牧草的营养品质较差是一个普遍现象。究其原因,土壤中营养元素含量较低,导致植物对氮、磷等元素的吸收利用效率较低。研究表明,氮素添加可迅速补充植物中的全氮含量,满足植物对营养元素的需求,进而恢复草地生产力,改善草地群落结构和植物的营养品质[25]。粗蛋白和粗脂肪含量是评价牧草营养价值的重要指标,其含量高则表明牧草营养品质较高,而粗纤维和粗灰分含量越高,则表明牧草可消化养分低,品质下降[26]。宋建超等[27]在高寒区的研究表明,氮添加显著提高了垂穗披碱草(Elymus nutans Griseb.)的粗蛋白和粗脂肪含量,与本研究结论一致。本研究中,所有施氮处理均显著提高了饲草的粗蛋白和粗脂肪含量。此外,我们还发现不同氮素形态和施氮水平对饲草的粗蛋白和粗脂肪含量具有显著影响,施用酰胺态氮肥更有利于粗蛋白和粗脂肪的积累,且高氮水平的促进作用更加明显。该现象一方面说明了高寒区氮限制非常严重,改善牧草的营养品质可能需要投入更多的氮肥;另一方面,由于不同的植物或生育期对氮肥的需求量和氮素形态具有明显差异,所以应根据实际情况选择适宜的氮素形态和施氮量。适宜的氮素形态是提高氮素利用率以及植物蛋白质含量的重要途径之一[28]。本研究中,酰胺态氮对植物粗蛋白和粗脂肪的促进效果明显高于铵态氮和硝态氮,这可能与植物的选择吸收能力以及不同氮肥的供氮能力等因素有关[29]。此外,本研究中的3种氮肥均不同程度地提高了饲草的粗灰分含量,降低了饲草的中性洗涤纤维及酸性洗涤纤维含量,与前人研究结论相似[30]。聚类分析将10个处理分为4大类,通过对比试验数据发现,这4大类可划分饲草生产性能和营养品质的优劣,基本对应了不施氮、低氮、中氮和高氮4种属性,因此,施氮水平对饲草生产性能和营养品质的影响是显而易见的。最后,本研究通过灰色关联度分析得出,选用酰胺态氮肥,施氮量为67.5 kg·hm−2·a−1时,多年生栽培草地饲草的生产性能和营养品质综合表现最优,研究结果可为青藏高原环青海湖地区人工草地生产力及营养品质的提高提供参考。
4. 结论
氮素形态和施氮水平对多年生高寒栽培草地饲草的生产性能和营养品质具有显著影响。选择酰胺态氮肥,施氮量为67.5 kg·hm−2·a−1时,多年生栽培草地饲草的生产性能和营养品质综合表现最优,说明适宜的氮肥管理制度对提高研究区饲草生产性能和营养价值具有促进作用。
-
表 1 额尔齐斯河流域河谷林种子植物物种组成及生活型
Table 1 Seed plant species composition and life form in valley forests of the Irtysh River basin
分类群Taxon 物种组成及占比Species composition and proportion / % 生活型Life form 科Family 属Genera 种Species 乔木Tree 灌木Shrub 草本Herb 藤本Vine 裸子植物 1 3 3 3 0 0 0 (1.82) (1.31) (0.74) 被子植物 单子叶植物 6 38 57 0 0 57 0 (10.91) (16.67) (14.14) 双子叶植物 48 187 343 18 52 267 6 (87.27) (82.02) (85.12) 总计 55 228 403 21 52 324 6  下载: 导出CSV
下载: 导出CSV
表 2 额尔齐斯河流域河谷林草本植物生活型统计
Table 2 Life form composition of herbaceous plants in valley forests of the Irtysh River basin
分类群Perennial herbs 种数No. of species 比例Proportion / % 多年生 222 68.52 一年生 61 18.83 一、二年生 23 7.10 二年生 11 3.40 一年或多年生 3 0.92 二年或多年生 3 0.92 一、二年生或多年生 1 0.31
下载: 导出CSV
表 3 额尔齐斯河流域河谷林种子植物科的分级统计
Table 3 Statistics on seed plant families in valley forests of the Irtysh River basin
级别
Grade科数
No. of families占总科比例
Proportion / %属数
No. of genera占总属比例
Proportion / %种数
No. of species占总种比例
Proportion / %含种数多的科(≥30种) 4 7.27 79 34.65 147 36.48 含种数较多的科(15~29种) 4 7.27 36 15.79 77 19.11 含种数一般的科(6~14种) 12 21.82 55 24.12 97 24.07 含种数较少的科(2~5种) 21 38.19 44 19.30 68 16.87 只含一个种的科(仅含1种) 14 25.45 14 6.14 14 3.47
下载: 导出CSV
表 4 额尔齐斯河流域河谷林种子植物属的分级统计
Table 4 Statistics on seed plant genera in valley forests of the Irtysh River basin
级别
Grade属数
No. of genera占总属比例
Proportion / %种数
No. of species占总种比例
Proportion / %含种数多的属(≥10种) 2 0.87 20 4.96 含种数一般的属(5~9种) 9 3.95 52 12.91 含种数较少的属(2~4种) 79 34.65 193 47.89 只含一个种的属(仅含1种) 138 60.53 138 34.24
下载: 导出CSV
表 5 额尔齐斯河流域河谷林种子植物科分布区类型
Table 5 Areal-types of seed plant families in valley forests of the Irtysh River basin
分布区类型Areal-type 河谷林Valley forests 新疆Xinjiang 科数
Family占比
Proportion / %科数
Family占比
Proportion / %1 世界广布 33 60.00 52 46.02 2 泛热带分布 8 14.55 17 15.04 2-2 热带亚洲-热带非洲-热带美洲分布 1 1.82 2 1.77 2S 以南半球为主的泛热带分布 − − 3 2.65 3 东亚(热带、亚热带)及热带南美间断 − − 3 2.65 8 北温带分布 4 7.27 12 10.62 8-4 北温带和南温带间断分布 7 12.73 14 12.39 8-5 欧亚和南美洲温带间断分布 1 1.82 2 1.77 9 东亚-北美间断分布 − − 1 0.88 10 旧世界温带分布 1 1.82 3 2.65 10-3 欧亚和南非(有时也在澳大利亚) − − 1 0.89 12-1 地中海区至中亚和南非洲和/或大洋洲间断分布 − − 1 0.89 12-5 温带至亚热带的盐土或沙漠和地中海地区 − − 1 0.89 13-2 中亚东部至喜马拉雅和中国西南部 − − 1 0.89 合计 55 100 113 100 热带性科/温带性科 0.692 3 0.694 4
下载: 导出CSV
表 6 额尔齐斯河流域河谷林种子植物属的分布区类型
Table 6 Areal-types of seed plant genera in valley forests of the Irtysh River basin
分布区类型Areal-type 河谷林Valley forests 新疆Xinjiang 属数Genera 占比Proportion / % 属数Genera 占比Proportion / % 1 世界广布 45 19.74 76 9.91 2 泛热带分布 7 3.07 39 5.08 2-1 热带亚洲-大洋洲和热带美洲分布 1 0.44 5 0.65 2-2 热带亚洲-热带非洲-热带美洲 - - 6 0.78 3 东亚(热带、亚热带)与热带美洲间断分布 1 0.44 5 0.65 4 旧世界热带分布 2 0.88 7 0.91 4-1 热带亚洲、非洲和大洋洲间断或星散分布 - - 2 0.26 5 热带亚洲及热带大洋洲 - - 4 0.52 6 热带亚洲至热带非洲 - - 4 0.52 7-1爪哇(或苏门答腊),喜马拉雅间断或星散分布到华南、西南 - - 1 0.13 7-4 越南(或中南半岛)至华南或西南分布 - - 1 0.13 7a 西马来,基本上在新华莱士线以西 - - 1 0.13 7a-c 西马来东马来间断分布 1 0.44 1 0.13 7a-d 西马来至东马来东达新几内亚 - - 1 0.13 7d 新几内亚分布 1 0.44 1 0.13 7e 热带亚洲,全分布区东南达西太平洋诸岛弧,包括新喀里多尼亚和斐济 - - 1 0.13 8 北温带分布 41 17.98 99 12.91 8-1 环极分布 1 0.44 2 0.26 8-2 北极-高山分布 1 0.44 12 1.56 8-4 北温带和南温带间断分布 49 21.49 104 13.56 8-5 欧亚和南美洲温带间断分布 6 2.63 17 2.22 9 东亚-北美间断分布 4 1.74 18 2.35 9-1墨西哥高山 - - 1 0.13 10 旧世界温带分布 27 11.84 91 11.86 10-1 地中海区,西亚(或中亚)和东亚间断分布 2 0.88 13 1.69 10-2 地中海区和喜马拉雅间断分布 3 1.32 5 0.65 10-3 欧亚和南非分布 9 3.95 19 2.48 11 温带亚洲分布 4 1.74 29 3.78 12 地中海区、西亚至中亚分布 16 7.02 80 10.43 12-1 地中海区至中亚和南非洲和/或大洋洲间断分布 1 0.44 8 1.04 12-2 地中海区至西亚或中亚和墨西哥或古巴间断 - - 1 0.13 12-3 地中海区至温带-热带亚洲,大洋洲和/或北美南部至南美洲间断分布 1 0.44 4 0.52 12-5 温带至亚热带的盐土或沙漠和地中海地区 - - 1 0.13 12-6 马加罗尼西亚,地中海欧洲,索科特拉,西亚,南至索马里分布 - - 1 0.13 13 中亚分布 1 0.44 45 5.87 13-1 中亚东部分布 2 0.88 8 1.04 13-2 中亚东部至喜马拉雅和中国西南部分布 1 0.44 19 2.48 13-3 西亚至喜马拉雅和西藏分布 - - 1 0.13 14 东亚 - - 5 0.65 14SH 中国-喜马拉雅分布 - - 10 1.30 14SJ 中国-日本分布 1 0.44 6 0.78 15 中国特有分布 - - 13 1.69 合计 228 100 767 100 热带性属/温带性属 0.076 5 0.129 1
下载: 导出CSV
表 7 额尔齐斯河流域河谷林重点保护野生植物名录
Table 7 List of key protected plants in valley forests of the Irtysh River basin
种名 Species 国家保护级别2021
Protected grade of
country自治区保护级别2024
Protected grade of
Xinjiang濒危等级2020
Endangered grade of
China’s Red List of Biodiversity额河杨 Populus×berolinensis var. jrtyschensis
(C. Y. Yang) C. ShangⅡ EN 新疆冷杉 Abies sibirica Korsh. Ⅰ EN 灰胡杨 Populus pruinose Schrenk Ⅱ 柔毛杨 Populus Pilosa Rehder EN 新疆落叶松 Larix sibirica Ledeb. VU 黑果枸杞 Lycium ruthenicum Murray Ⅱ 新疆枸杞 Lycium dasystemum Pojark. Ⅱ 额河木蓼 Atraphaxis irtyschensis Chang Y. Yang & Y. L. Han Ⅱ VU 越桔 Vaccinium vitis-idaea L. Ⅱ 新疆百合 Lilium martagon var. pilosiusculum Freyn Ⅱ 甘草 Glycyrrhiza uralensis Fisch. Ⅱ 阿尔泰葱 Allium altaicum Pall. Ⅱ 大赖草 Leymus racemosus (Lam.) Tzvelev Ⅱ 红景天 Rhodiola rosea L. Ⅱ VU 块根芍药 Paeonia anomala C. A. Mey. Ⅰ VU 大叶补血草 Limonium gmelinii (Willd.) Kuntze VU 巨车前 Plantago maxima Juss. ex Jacq. VU 裸花蜀葵 Althaea nudiflora (Lindl.) Boiss. EN 裂唇虎舌兰 Epipogium aphyllum (F. W. Schmidt) Sw. Ⅰ EN 紫斑掌裂兰 Dactylorhiza fuchsia (Druce) Soó Ⅰ 紫点掌裂兰 Dactylorhiza incarnata subsp. cruenta
(O. F. Müll.) P. D. SellⅠ 小斑叶兰 Goodyera repens (L.) R. Br. Ⅱ 新疆火烧兰 Epipactis palustris (L.) Crantz Ⅱ VU 阴生掌裂兰 Dactylorhiza umbrosa (Kar. & Kir.) Nevski Ⅱ 黄花软紫草 Arnebia guttata Bunge VU
下载: 导出CSV
-
[1] Daru BH,Elliott TL,Park DS,Jonathan Davies T. Understanding the processes underpinning patterns of phylogenetic regionalization[J]. Trends Ecol Evol,2017,32(11):845−860. doi: 10.1016/j.tree.2017.08.013
[2] Li R,Qian LS,Sun H. Current progress and future prospects in phylofloristics[J]. Plant Divers,2018,40(4):141−146. doi: 10.1016/j.pld.2018.07.003
[3] 向晓媚,刘冰,谭璐,陈功锡. 湖南省德夯风景名胜区种子植物区系研究[J]. 西北植物学报,2022,42(6):1051−1063. Xiang XM,Liu B,Tan L,Chen GX. Study on the flora of seed plants in Dehang scenic spot,Hunan Province[J]. Acta Botanica Boreali-Occidentalia Sinica,2022,42(6):1051−1063.
[4] 刘平,王健. 新疆额尔齐斯河河谷林经营现状及保护对策[J]. 林业资源管理,2002(1):35−37. Liu P,Wang J. Management and conservation of the forests in the Erchis River Valley in Xinjiang[J]. Forest and Grassland Resources Research,2002(1):35−37.
[5] 臧润国,张炜银,成克武,白志强,张新平,等. 新疆额尔齐斯河河谷天然杨树林群落特征与保育对策[C]//中国生物多样性保护与研究进展Ⅶ——第七届全国生物多样性保护与持续利用研讨会论文集. 长春:国际生物多样性计划中国委员会,2006:296−303. [6] 刘时栋,刘琳,张建军,王柯,郭义强. 基于生态系统服务能力提升的干旱区生态保护与修复研究——以额尔齐斯河流域生态保护与修复试点工程区为例[J]. 生态学报,2019,39(23):8998−9007. Liu SD,Liu L,Zhang JJ,Wang K,Guo YQ. Study on ecological protection and restoration path of arid area based on improvement of ecosystem service capability,a case of the ecological protection and restoration pilot project area in Irtysh River Basin[J]. Acta Ecologica Sinica,2019,39(23):8998−9007.
[7] Liu HC,Fan J,Liu BY,Wang L,Qiao Q. Practical exploration of ecological restoration and management of the mountains-rivers-forests-farmlands-lakes-grasslands system in the Irtysh River Basin in Altay,Xinjiang[J]. Journal of Resources and Ecology,2021,12(6):766−776.
[8] An SQ,Cheng XL,Sun SC,Wang YJ,Li J. Composition change and vegetation degradation of riparian forests in the Altai Plain,NW China[J]. Plant Ecol,2003,164:75−84. doi: 10.1023/A:1021225204808
[9] 努尔加那提·拉扎提. 阿勒泰地区生态环境与经济协调发展研究[D]. 乌鲁木齐:新疆师范大学,2014:11−38. [10] Qin Y,Kayrat A,Zhang Z,Liu P,Liu H. The main problems of the water ecological environment and protective countermeasures in the river basin of the Altay Region,Xinjiang[J]. Journal of Resources and Ecology,2023,14(2):383−390.
[11] 刘浩,郝海广,张文国,刘鹏,孙丽慧. 基于格局-质量-服务的生态保护修复成效评估:以额尔齐斯河流域为例[J]. 环境科学研究,2022,35(11):2495−2507. Liu H,Hao HG,Zhang WG,Liu P,Sun LH. Evaluation of ecological protection and restoration effectiveness based on ‘pattern-quality-service’ in Irtysh River Basin[J]. Research of Environmental Sciences,2022,35(11):2495−2507.
[12] 王希群,郭保香,张利. 新疆额尔齐斯河科克托海湿地自然保护区保护价值[J]. 林业资源管理,2016(5):6−12. Wang XQ,Guo BX,Zhang L. Protection values on Xinjiang Irtysh Keketuohai wetland nature reserve[J]. Forest Resources Management,2016(5):6−12.
[13] 新疆荒地资源综合考察队. 新疆重点地区荒地资源合理利用[M]. 乌鲁木齐:新疆人民出版社,1985:1−326. [14] 吴征镒,周浙昆,孙航,李德铢,彭华. 种子植物分布区类型及其起源和分化[M]. 昆明:云南科技出版社,2006:1−451. [15] 李德铢,陈之瑞,王红,路安民. 中国维管植物科属词典[M]. 北京:科学出版社,2018:1−682. [16] 黄继红,马克平,陈彬. 中国特有种子植物的多样性及其地理分布[M]. 北京:高等教育出版社,2014:1−100. [17] 张天汉,王健铭,夏延国,郑昌龙,李景文,等. 库姆塔格沙漠不同区域种子植物物种组成与区系特征研究[J]. 植物科学学报,2016,34(1):78−88. Zhang TH,Wang JM,Xia YG,Zheng CL,Li JW,et al. Composition of seed plant species and flora features in different areas of the Kumtag Desert of China[J]. Plant Science Journal,2016,34(1):78−88.
[18] 张学渊,魏伟,周亮,郭泽呈,李振亚,等. 西北干旱区生态脆弱性时空演变分析[J]. 生态学报,2021,41(12):4707−4719. Zhang XY,Wei W,Zhou L,Guo ZC,Li ZY,et al. Analysis on spatio-temporal evolution of ecological vulnerability in arid areas of Northwest China[J]. Acta Ecologica Sinica,2021,41(12):4707−4719.
[19] 中国植被编辑委员会. 中国植被[M]. 北京:科学出版社,1980:1−1391. [20] 张继义,赵哈林,张铜会,赵学勇. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态[J]. 植物生态学报,2004,28(1):86−92. Zhang JY,Zhao HL,Zhang TH,Zhao XY. Dynamics of species diversity of communities in restoration processes in Horqin Sandy Land[J]. Acta Phytoecologica Sinica,2004,28(1):86−92.
[21] 欧尔辉,周鸿玉,杨秀玲,魏民,邓磊. 共和盆地东缘人工植被演替序列的群落物种多样性格局[J]. 青海大学学报,2023,41(1):1−7. Ou EH,Zhou HY,Yang XL,Wei M,Deng L. Community species diversity pattern of artificial vegetation succession sequences in the eastern margin of Gonghe Basin[J]. Journal of Qinghai University,2023,41(1):1−7.
[22] 王雅芳,李国旗,石云,刘星,谢博勋,解盛. 贺兰山低山区土壤种子库与地上植被关系分析[J]. 草地学报,2023,31(1):166−172. Wang YF,Li GQ,Shi Y,Liu X,Xie BX,Xie S. Analysis of relationship between soil seed bank and aboveground vegetation in low mountain area of Helan Mountain[J]. Acta Agrestia Sinica,2023,31(1):166−172.
[23] 闫建成,梁存柱,付晓玥,王炜,王立新,贾成朕. 草原与荒漠一年生植物性状对降水变化的响应[J]. 草业学报,2013,22(1):68−76. Yan JC,Liang CZ,Fu XY,Wang W,Wang LX,Jia CZ. The responses of annual plant traits to rainfall variation in steppe and desert regions[J]. Acta Prataculturae Sinica,2013,22(1):68−76.
[24] 席璐璐,缑倩倩,王国华,宋冰. 荒漠绿洲过渡带一年生草本植物对干旱胁迫的响应[J]. 生态学报,2021,41(13):5425−5434. Xi LL,Gou QQ,Wang GH,Song B. The responses of typical annual herbaceous plants to drought stress in a desert-oasis ecotone[J]. Acta Ecologica Sinica,2021,41(13):5425−5434.
[25] 任亦君,席璐璐,缑倩倩,宋冰,王国华. 单次小降雨( ≤ 5 mm)事件对4种典型荒漠一年生草本植物生长和繁殖的影响[J]. 中国沙漠,2021,41(4):87−99. Ren YJ,Xi LL,Gou QQ,Song B,Wang GH. Effects of single small rainfall events ( ≤ 5 mm) on the growth and reproduction of four typical desert annual herbaceous plants[J]. Journal of Desert Research,2021,41(4):87−99.
[26] 崔乃然,李学禹. 新疆极端环境条件下的植物种质资源[J]. 石河子大学学报(自然科学版),1998,2(4):48−63. Cui NR,Li XY. The plant plasm resources under extreme enviromental conditions in Xinjiang[J]. Journal of Shihezi University (Natural Science),1998,2(4):48−63.
[27] 陈文俐,杨昌友. 中国阿尔泰山种子植物区系研究[J]. 云南植物研究,2000,22(4):371−378. Chen WL,Yang CY. A floristic study on the seed plant in Mts. Altay of China[J]. Acta Botanica Yunnanica,2000,22(4):371−378.
[28] 秦仁昌. 新疆阿尔泰山的植物区系、植被类型和植物资源[J]. 科学通报,1957,8(3):114−115. [29] 曹秋梅,尹林克,陈艳锋,杨美琳,杨更强. 阿尔泰山南坡种子植物区系特点分析[J]. 西北植物学报,2015,35(7):1460−1469. Cao QM,Yin LK,Chen YF,Yang ML,Yang G Q. Analysis on characteristics of flora in south slope of Altai Mountain[J]. Acta Botanica Boreali-Occidentalia Sinica,2015,35(7):1460−1469.
[30] 韩路,陈家力,王家强,王海珍,吕瑞恒,康佳鹏. 塔河源荒漠河岸林群落物种组成、结构与植物区系特征[J]. 植物科学学报,2019,37(3):324−336. Han L,Chen JL,Wang JQ,Wang HZ,Lü RH,Kang JP. Species composition,community structure,and floristic characteristics of desert riparian forest community along the mainstream of Tarim River[J]. Plant Science Journal,2019,37(3):324−336.
[31] 孔晓晶. 准噶尔盆地荒漠植物区系及植被图绘制的初步研究[D]. 乌鲁木齐:新疆农业大学,2019:1−38. [32] 徐远杰,陈亚宁,李卫红,孙慧兰,陈亚鹏. 中国伊犁河谷种子植物区系分析[J]. 干旱区研究,2010,27(3):331−337. Xu YJ,Chen YN,Li WH,Sun HL,Chen YP. Analysis on the spermatophyte floras in the Ili River valley of China[J]. Arid Zone Research,2010,27(3):331−337.
[33] 余若云,黄继红,郭仲军,唐素英,王立平,等. 新疆种子植物区系的数量统计及其地理成分特征[J]. 世界生态学,2018,7(3):193−205. doi: 10.12677/IJE.2018.73021 Yu RY,Huang JH,Guo ZJ,Tang SY,Wang LP,et al. Quantitative statistics and geographical components of flora of seed plants in Xinjiang[J]. International Journal of Ecology,2018,7(3):193−205. doi: 10.12677/IJE.2018.73021
[34] 潘晓玲,张宏达. 准噶尔盆地植被特点与植物区系形成的探讨[J]. 中山大学学报论丛,1996(2):97−101. Pan XL,Zhang HD. Research on the characteristics of vegetation and the forming of flora in Zhungeer Basin,Xinjiang[J]. Supplement to the Journal of Sun Yatsen University,1996(2):97−101.
[35] 于军,王海珍,陈加利,韩路. 塔里木河流域荒漠河岸林胡杨群落的空间格局研究[J]. 中国沙漠,2011,31(4):913−918. Yu J,Wang HZ,Chen JL,Han L. Spatial pattern of Populus euphratica community of desert riparian forest in Tarim River Basin[J]. Journal of Desert Research,2011,31(4):913−918.
[36] 张明理. 中国西北干旱区和中亚植物区系地理研究[J]. 生物多样性,2017,25(2):147−155. Zhang ML. A review on the floristic phytogeography in arid Northwestern China and central Asia[J]. Biodiversity Science,2017,25(2):147−155.
[37] Miao YF,Meng QQ,Fang XM,Yan XL,Wu FL,Song CH. Origin and development of Artemisia (Asteraceae) in Asia and its implications for the uplift history of the Tibetan Plateau:a review[J]. Quat Int,2011,236(1-2):3−12. doi: 10.1016/j.quaint.2010.08.014
[38] Zhang ML,Hao XL,Sanderson SC,Vyacheslav BV,Sukhorukov AP,Zhang X. Spatiotemporal evolution of Reaumuria (Tamaricaceae) in central Asia:insights from molecular biogeography[J]. Phytotaxa,2014,a,167(1):89−103.
[39] Zhang ML,Sanderson SC,Sun YX,Byalt VV,Hao XL. Tertiary montane origin of the central Asian flora,evidence inferred from cpDNA sequences of Atraphaxis (Polygonaceae)[J]. J Integr Plant Biol,2014,b,56(12):1125−1135.
[40] 潘晓玲,张宏达. 哈纳斯自然保护区植被特点及植物区系形成的探讨[J]. 干旱区研究,1994,11(4):1−7. Pan XL,Zhang HD. Character of vegetation and research on forming of flora in Kanas[J]. Arid Zone Research,1994,11(4):1−7.
[41] 党荣理,潘晓玲. 西北干旱荒漠区植物区系的特有现象分析[J]. 植物研究,2001,21(4):519−526. Dang RL,Pan XL. The Chinese endemic plant analysis in west-north desert of China[J]. Bulletin of Botanical Research,2001,21(4):519−526.
[42] 成克武. 新疆额尔齐斯河流域天然林保育技术研究[D]. 北京:中国林业科学研究院,2005:9−126. [43] 刘海丰,李亮,桑卫国. 东灵山暖温带落叶阔叶次生林动态监测样地:物种组成与群落结构[J]. 生物多样性,2011,19(2):232−242. doi: 10.3724/SP.J.1003.2011.11319 Liu HF,Li L,Sang WG. Species composition and community structure of the Donglingshan forest dynamic plot in a warm temperate deciduous broad-leaved secondary forest,China[J]. Biodiversity Science,2011,19(2):232−242. doi: 10.3724/SP.J.1003.2011.11319
计量
- 文章访问数: 396
- HTML全文浏览量: 93
- PDF下载量: 24
