
Construction and evaluation of Polygonatum cyrtonema Hua intercropping based on the growth and physiological adaptability
-
摘要:
多花黄精(Polygonatum cyrtonema Hua)多采用林下仿野生种植,猕猴桃(Actinidia chinensis Planch.)果园林下环境与森林接近,具备复合种植的潜在可行性。本研究通过设置不同猕猴桃品种(‘红阳’、‘金艳’)、郁闭度(60%、30%、全光照)和种植密度(株行距25 cm×25 cm、25 cm×40 cm、35 cm×40 cm),分析对多花黄精生长的影响,以筛选适宜的种植模式。结果显示:‘金艳’果园中,多花黄精植株高度较高,基径较粗;60%郁闭度果园中,多花黄精叶片叶绿素含量较高,丙二醛(MDA)含量和抗坏血酸过氧化物酶(APX)活性较低;30%郁闭度果园中,植株高度较高;而全光照条件下,叶片相对含水量较低,可溶性蛋白、脯氨酸、MDA、总酚、类黄酮含量较高,过氧化氢酶(CAT)和APX活性较高;低等密度模式中叶片MDA和总酚含量较低,而高等密度中叶片MDA含量较高。综合而言,猕猴桃果园复合种植多花黄精是可行的,且适宜的种植模式为郁闭度30%~60%、种植株行距(25~35) cm×40 cm。
Abstract:Polygonatum cyrtonema Hua is typically cultivated under forest conditions that mimic its natural habitat. The understory environment of Actinidia chinensis Planch. orchards shares similar ecological characteristics with forests, suggesting the potential feasibility of intercropping P. cyrtonema within A. chinensis orchards. However, optimizing an appropriate planting model and creating a suitable growth environment for P. cyrtonema requires experimental validation. This study assessed the growth and physiological adaptability of P. cyrtonema under different intercropping conditions by varying A. chinensis cultivars (‘Hongyang’ and ‘Jinyan’), canopy densities (60%, 30%, and full sunlight), and planting densities (row spacing of 25 cm×25 cm, 25 cm×40 cm, and 35 cm×40 cm). Results demonstrated that P. cyrtonema exhibited greater plant height and basal diameter in orchards planted with A. chinensis cv. Jinyan. Under 60% canopy density, chlorophyll content was elevated, while malondialdehyde (MDA) content and ascorbate peroxidase (APX) activity were lower. A 30% canopy density promoted greater plant height. In full sunlight, leaves had reduced relative water content but exhibited increased soluble protein, proline, MDA, total phenol, and flavonoid concentrations, along with higher catalase (CAT) and APX activity. Lower planting densities resulted in reduced MDA and total phenol concentrations in leaves, whereas higher planting densities were associated with increased MDA content. These findings indicate that intercropping P. cyrtonema within A. chinensis orchards is feasible. Optimal planting conditions were a canopy density of 30%–60% and a row spacing of (25–35) cm×40 cm.
-
植物群落中物种数目随样地面积的增加而增加,这已经成为“植物群落生态学中的定律之一”[1]。随着取样面积的扩大,样方内的种类也随之增加,最初增长很快,以后逐渐缓慢,形成一条曲线,叫做种-面积曲线(Species-area curve)[2]。对野外调查获得的种-面积数据进行曲线模型拟合,还可以进一步确定植物群落物种多样性调查的最小取样面积[3]。最小面积是反映群落种类组成和结构特征的最小地段[4]。最小取样面积的确认有助于制定合理的最小取样面积,同时可以降低昂贵的调查成本,对生态学研究具有重要的意义。最常用的3类种-面积关系拟合模型为对数函数模型(Logarithmic function)、幂函数模型(Power function)和逻辑斯蒂模型(Logistic function)[5-9]。Scheiner[10]发现种-面积关系的构建方式包括组合样方法、巢式样方法和隔离生境。众多研究表明,不同群落所需的最小取样面积明显不同[11-13]。不同群落的灌、草层会受到不同乔木层特征差异的影响,因此在研究中,不同学者也习惯将一个群落分为乔、灌、草3层(或者更多)分别进行拟合[14-17]。
海岛由于受风害的干扰,以及自身土壤的贫瘠性,生态环境十分的敏感和脆弱。国内学者对海岛森林的群落特征、多样性、碳储量等方面做了许多的研究[18-20],部分学者研究了物种数和岛屿面积的关系,如王金旺等[21]对温州沿海20个小型无居民海岛进行了种-面积关系拟合,结果表明种-面积关系不存在小岛屿效应,岛屿植物物种丰富度主要受面积影响,但又不总是由岛屿面积决定。肖兰等[22]对厦门无居民海岛植物物种数和海岛面积关系进行了研究,结果表明物种丰富度受海岛面积的影响较大,与海岛面积存在显著的对数和幂函数关系。上述研究是针对海岛总物种数和海岛总面积的关系,其并未研究物种数随着面积扩大而在该岛屿上发生的变化情况,目前也未见针对不同群落在海岛上的种-面积关系和最小取样面积的相关报道。福州市计划将琅岐岛建设成为生态旅游岛,但琅岐岛土壤十分贫瘠,渗漏性强,保蓄力弱,养分含量甚低,同时风沙灾害频发,植物的生存环境恶劣[23]。且随着海岛的不断开发与建设,特别是海岛人工林的建设,亟待研究不同群落的物种组成和多样性特征等相关数据,为生态建设提供参考。海岛与内陆环境存在较大差异,目前内陆森林相关报道对海岛森林相关研究的借鉴意义不大,而通过对琅岐岛主要森林群落种-面积关系进行分析,能给琅岐岛未来的开发建设提供一定的参考。
种-面积曲线只考虑物种种类的增加,受各种因素的影响,在实际工作中能代表整个群落组成结构特征的最小面积几乎不存在,只能尽量增加取样面积以减小误差,但是这又会导致工作成本的增加。此前植物多样性的调查大多是根据经验值确定调查样方的面积,比如常用的亚热带森林为20 m × 20 m[24]。这种根据经验来设定样方面积,并不完全适用于所有群落调查。因为在研究一个群落时,对该群落进行全部调查会浪费大量的调查成本且难以实现。因此,从植物的保护、修复和海岛开发和利用等角度综合考虑,对琅岐岛不同森林群落物种多样性调查的最小取样面积研究不仅对保护该地区的物种和生物多样性具有重要作用,同时也有利于以后学者在进行相关研究时能选择适合的样方面积,提高工作效率,减少调研成本。
位于琅岐岛云龙村旁风成沙丘上的天然朴树(Celtis sinensis Pers.)林,距今已有400多年历史;而岛上的台湾相思(Acacia confusa Merr.)林和柠檬桉林(Eucalyptus citriodora Hook. f.)为20世纪飞机撒播造林或人工造林形成,人为影响和干预较小,属于半自然群落。3类群落在当地占地面积和分布十分广泛,是琅岐岛主要的组成树种,本研究在综合分析琅岐岛这3类主要森林群落的实际情况后,先根据亚热带森林经验取样20 m × 20 m作为样地样方面积,采用巢式样方法[25]进行3类森林群落的种-面积曲线调查。利用3种模型分别拟合整体群落、灌木(包括木质藤本及胸径 ≤ 3 cm且树高 ≤ 3 m的乔木幼树)和草本(包括草质藤本和蕨类)3层的方程(由于海岛群落乔木层种类较少,因此不单独分析),并通过拟合优度检验,筛选最优方程,利用不同估测精度要求和拐点法进行最小取样面积的计算,得到相应的最小取样面积,以期为后续琅岐岛的森林多样性调查研究提供一定的参考依据。
1. 材料与方法
1.1 研究区概况
福州琅岐岛位于闽江入海口,全岛总面积约92 km2,其中陆地面积约55 km2(平原约36.5 km2,山地约18.5 km2),全岛森林覆盖率达20%。岛东西长约15.3 km,南北宽约8.1 km。岛上土壤盐渍化严重,风沙灾害频发,植物的生存环境恶劣,岛上主要优势树种包括木麻黄(Casuarina equisetifolia L.)、台湾相思、柠檬桉、朴树和高山榕(Ficus altissima Blume)[26]。经现场调研发现,琅岐岛上台湾相思、柠檬桉、朴树3种森林群落呈片状、块状分布;木麻黄林作为沿海地区防护林,基本为带状分布,且基本为幼、中林龄;高山榕多为混交或分布于人工群落。
1.2 取样方法
本研究基于海岛地形、地图影像、森林群落分布及可达性等因素,采用典型取样法,在干扰小,距离林缘大于30 m,林龄达成熟林的区域选择具有代表性的群落。本研究选择了台湾相思林、柠檬桉林、朴树林共3类森林群落的18个20 m × 20 m的样地,每个样地4次重复(即从4个角分别扩大面积记录物种数,并取平均值进行计算),则共计72个(每种森林群落24个)20 m × 20 m的样方。其中朴树林由于集中分布在云龙村旁,因此6个样地均位于此,海拔为11.09 ~ 35.27 m,坡度为16° ~ 37°,该样地占整个琅岐岛朴树林面积80%左右,台湾相思林和柠檬桉林则分散于琅岐岛各处(表1,附表1
1 )。每个样地植物丰富、生长均匀、郁闭度高于0.75,且面积满足20 m × 20 m的森林群落进行种-面积调查。野外调查方法为相邻样方格子法[27],在每个样方左上角开始,以1 m为间隔纵横拉线,于是20 m × 20 m的大样方就被分割为包含400个方格的网格样方,每个网格样方面积为1 m2[28],方便后续记录,再按照巢式样方法开展物种丰富度的调查,即在样地的4个角的第一个1 m2小样方作为第一个面积梯度,然后逐步扩大调查样方面积,记录每个样方内新出现的物种名称、生活类型,直至覆盖整个样地[25]。本研究的群落中,首先设置1 m × 1 m的样方为1,作为第一个调查样方;将1、2合并,作为第二个调查样方;1、2、3合并……;则整体森林群落的调查样方面积依次为1 m × 1 m、1 m × 2 m、2 m × 2 m、2 m × 4 m、……、8 m × 16 m、16 m × 16 m、16 m × 20 m、20 m × 20 m(即每个样方11个面积梯度,扩大方式如图1所示)。表 1 样地基础信息Table 1. Basic information of sample plots样地号
Sample plot number海拔
Altitude / m坡向
Aspect / °坡度
Slope / °总物种数量
Total number of species总灌木数量
Total number of shrubs总草本数量
Total number of herbs台湾相思
Acacia confusa Merr.1 15.11 西北 NW287 12 24 14 10 2 10.37 西北 NW312 25 59 36 22 3 158.57 西南 SW244 34 67 46 19 4 118.76 东北 NE20 29 78 46 32 5 31.57 东南 SE158 16 57 37 20 6 14.39 东南 SE144 27 43 28 15 柠檬桉
Eucalyptus citriodora
Hook. f.1 18.14 西南 SW220 10 51 27 23 2 5.67 西南 SW259 23 69 40 28 3 78.40 西南 SW206 34 82 52 27 4 61.34 西 W279 15 58 25 33 5 24.10 东南 SE169 11 42 20 19 6 103.89 东北 NE68 31 74 50 24 朴树
Celtis sinensis Pers.1 35.27 南 S188 37 43 30 14 2 25.43 东南 SE143 25 41 25 16 3 25.43 东南 SE130 22 32 22 9 4 25.43 西 W270 30 48 31 17 5 25.43 西南 SW244 33 30 17 13 6 11.09 东南 SE162 35 30 20 10 ![]() 图 1 巢式样方法合并扩大示意图巢氏样方面积是按照2倍增加扩散,直至覆盖全部样方。1代表巢式样方法的第1个取样面积,面积为1 m × 1 m;1和2合并后代表巢式样方法的第2个取样面积,面积为1 m × 2 m;1、2和3合并后代表巢式样方法的第3个取样面积,面积为2 m × 2 m;……;1 ~ 11合并后代表巢式样方法的第11个取样面积,面积为20 m × 20 m。Figure 1. Schematic of nested method of merging expansion1 Sampling area of the nested sample is increased by two times until it covers all samples. 1 represents the first sampling area of the nested sampling method, with an area of 1 m × 1 m. Combination of 1 and 2 represent the second sampling area of the nested sampling method, with an area of 1 m × 2 m. Combination of 1, 2, and 3 represent the third sampling area of the nested sampling method, with an area of 2 m × 2 m…… ; combination of 1–11 represents the 11th sampling area of the nested sampling method, with an area of 20 m × 20 m.
图 1 巢式样方法合并扩大示意图巢氏样方面积是按照2倍增加扩散,直至覆盖全部样方。1代表巢式样方法的第1个取样面积,面积为1 m × 1 m;1和2合并后代表巢式样方法的第2个取样面积,面积为1 m × 2 m;1、2和3合并后代表巢式样方法的第3个取样面积,面积为2 m × 2 m;……;1 ~ 11合并后代表巢式样方法的第11个取样面积,面积为20 m × 20 m。Figure 1. Schematic of nested method of merging expansion1 Sampling area of the nested sample is increased by two times until it covers all samples. 1 represents the first sampling area of the nested sampling method, with an area of 1 m × 1 m. Combination of 1 and 2 represent the second sampling area of the nested sampling method, with an area of 1 m × 2 m. Combination of 1, 2, and 3 represent the third sampling area of the nested sampling method, with an area of 2 m × 2 m…… ; combination of 1–11 represents the 11th sampling area of the nested sampling method, with an area of 20 m × 20 m.1.3 拟合模型及对应的最小取样面积方程
本研究统计3类森林共18个样地、72个样方中,整体群落、灌木层和草本层3个对象分别在每个样方中的11个面积梯度的平均物种总数、平均灌木总数、平均草本总数的数据(每种森林类型3组,共9组数据),选择巢式样方法最常用的3个种-面积拟合模型及其对应的最小取样面积方程进行函数模型的拟合及相关计算(表2)。
表 2 巢式样方法常用的种-面积关系拟合模型Table 2. Common species-area relationship fitting models used in nested sampling method函数
Function种-面积拟合函数模型
Species-area fitting
function model最小取样面积方程
Minimum sampling
area equation对数 (1)S=a + blnA (4)A=exp[(ρSt−a)/b] 幂函数 (2)S=aAb (5)A=(ρSt / a)1/b 逻辑斯蒂 (3)S=c/(1 + ae−bA) (6)A=-{ln[(c/ρSt−1)/a]}/b 注:A表示样地面积;S表示对应样地的物种数目;a、b、c为待估参数;St表示调查群落物种总数;ρ表示期望物种与总物种的比例。 Notes: A represents sample plot area; S represents number of species in the corresponding sample plot; a, b, and c are parameters to be estimated; St represents total number of species in the surveyed community; ρ represents the ratio of expected species to total species. 1.4 种-面积关系拟合优度评价及最小取样面积确定
采用最小二乘法对对数模型进行检验,用高斯牛顿算法对幂函数模型及逻辑斯蒂模型进行检验[5]。采用赤池信息准则(Akaike information criterion,AIC)对3种模型的拟合优度进行评价[29]。AIC数值越小,模型拟合越好,预测值越准确。
本研究用两种方法结合分析最小样方面积。Archibald[30]研究发现,在占群落总物种数50%对应的面积上常见种就总是出现,因此将这个面积定义为群落最小面积。因此本研究选择5种不同的估测精度(50%、60%、70%、80%和90%)分别计算3类海岛森林群落3个对象的样方面积,并分析要得到群落总数一定比例(不同精度)的物种数所需最小面积差异[25, 31, 32]。同时基于最优模型采用拐点法计算最小调查取样面积[3, 32-34]。本研究拐点的具体步骤为:计算最优拟合模型的一阶导数,将不同精度下的最小取样面积A带入一阶导数,得到面积对应曲线的斜率;参考已有研究的做法,以曲线斜率 < 0.1作为识别拐点的标准[32, 34],进而确定最小取样面积。
本研究的数据分析中,逻辑斯蒂函数中的c值是利用Mathematica 12.2软件编程处理获得[35],AIC值是通过R 3.5.1软件编程计算,其他所有数据分析、作图采用R 3.5.1、SPSS 23、Excel 2019软件完成。
2. 结果与分析
2.1 研究地样方物种组成
本研究根据18个样地的调查结果,共记录物种数为219种,隶属于80科174属。其中禾本科植物种类最多,为16种,占总数的7.31%;其次为豆科14种,占总数的6.39%;菊科13种,占总数的5.94%(图2)。按照《中国植物志》和《福建植物志》的描述进行生活型分类,调查的18个样地中,乔木植物占比极小,共12种(占比5.29%),主要有台湾相思、朴树和柠檬桉;灌木101种(占比44.49%),主要有马缨丹(Lantana camara L.)、杠香藤(Mallotus repandus var. Chrysocarpus (Pamp.) S. M. Hwang)、蓬藟(Rubus hirsutus Thunb.)和福建胡颓子(Elaeagnus oldhamii Maximowicz)等;草本植物96种(占比42.29%),主要有荩草(Arthraxon hispidus (Trin.) Makino)、求米草(Oplismenus undulatifolius (Arduino) Beauv.)、积雪草(Centella asiatica (L.) Urban)和火炭母(Polygonum chinense (L.) H. Gross)等;藤本植物18种(占比7.93%),主要有细圆藤(Pericampylus glaucus (Lam.) Merr.)、山葛(Pueraria montana (Loureiro) Merrill)和匙羹藤(Gymnema sylvestre (Retz.) Schult.)(图3)。台湾相思群落平均每个样方物种数为54.7种、灌木34.5种和草本19.7种;柠檬桉群落平均每个样方物种数有62.7种、灌木35.7种和草本25.7种;朴树群落平均每个样方物种数有37.3种、灌木24.2种和草本13.2种。
![]() 图 2 主要科的物种数1:禾本科;2:豆科;3:菊科;4:大戟科;5:蔷薇科;6:百合科;7:马鞭草科。Figure 2. Number of species in major families1: Gramineae; 2: Fabaceae; 3: Asteraceae; 4: Euphorbiaceae; 5: Rosaceae; 6: Liliaceae; 7: Verbenaceae.
图 2 主要科的物种数1:禾本科;2:豆科;3:菊科;4:大戟科;5:蔷薇科;6:百合科;7:马鞭草科。Figure 2. Number of species in major families1: Gramineae; 2: Fabaceae; 3: Asteraceae; 4: Euphorbiaceae; 5: Rosaceae; 6: Liliaceae; 7: Verbenaceae.2.2 各群落3个对象植物种-面积关系的最优拟合模型
利用3类海岛森林群落的野外调查数据对函数(1) ~ (3)进行拟合,求解得到相应的种-面积曲线方程的参数,从R2(R2 > 0.9,P < 0.01)发现3类海岛森林群落3个对象植物的种-面积关系都符合3种拟合函数模型(表3、图4、图5),进一步通过赤池信息准则(AIC)对3种函数模型的拟合优度进行评价,从而可以确定相应的最优种-面积曲线(表3)。3类森林群落整体的幂函数模型拟合效果最好(AIC值分别为−24.429、−36.845、−37.642),其次是逻辑斯蒂函数模型,对数函数模型的拟合效果最差;3类森林群落最优拟合结果的种-面积曲线见图4。3类森林群落灌木层(包括木质藤本)的幂函数模型拟合效果最好(AIC值分别为−19.093、−25.709、−39.172),其次是逻辑斯蒂函数模型,对数函数模型的拟合效果最差;3类森林群落灌木层最优拟合结果的种-面积曲线见图5。3类森林群落草本层(包括草质藤本和蕨类)的幂函数模型拟合效果最好(AIC值分别为−26.498、−38.749、−30.644),其次是逻辑斯蒂函数模型,对数函数模型的拟合效果最差(朴树的草本层对数函数较逻辑斯蒂函数模型拟合效果稍好);3类森林群落草本层最优拟合结果的种-面积曲线见图6。因此,台湾相思林、柠檬桉林、朴树林的种-面积关系最优拟合模型分别为:S台=11.813A0.269、S柠=10.642A0.307、S朴=7.774A0.271;灌木层的种-面积关系最优拟合模型分别为:S台灌=5.804A0.314、S柠灌=6.069A0.309、S朴灌=3.992A0.309;草本层的种-面积关系最优拟合模型分别为:S台草=6.121A0.206、S柠草=4.427A0.302、S朴草=3.712A0.223。
表 3 种-面积关系拟合结果Table 3. Fitting results of species-area relationship研究对象
Object of study森林类型
Forest type函数
Function参数 Parameter R² P AIC a b c 群落整体 台湾相思林 (1) 7.415 7.715 − 0.982 <0.01 53.716 (2) 11.813 0.269 − 0.988 <0.01 −24.429 (3) 0.034 0.986 55.00 0.938 <0.01 21.460 柠檬桉林 (1) 4.264 9.295 − 0.964 <0.01 65.706 (2) 10.642 0.307 − 0.997 <0.01 −36.845 (3) 0.420 0.984 62.92 0.973 <0.01 15.505 朴树林 (1) 4.521 5.204 − 0.973 <0.01 49.605 (2) 7.774 0.271 − 0.996 <0.01 −37.642 (3) 0.056 0.986 37.51 0.964 <0.01 15.897 灌木层 台湾相思林 (1) 2.511 5.188 − 0.978 <0.01 47.294 (2) 5.804 0.314 − 0.985 <0.01 −19.093 (3) 0.068 0.986 34.68 0.929 <0.01 24.050 柠檬桉林 (1) 2.614 5.316 − 0.972 <0.01 50.547 (2) 6.069 0.309 − 0.992 <0.01 −25.709 (3) 0.072 0.984 35.82 0.968 <0.01 17.431 朴树林 (1) 1.467 3.573 − 0.958 <0.01 46.131 (2) 3.992 0.309 − 0.997 <0.01 −37.172 (3) 0.111 0.985 24.34 0.962 <0.01 17.406 草本层 台湾相思林 (1) 5.043 2.444 − 0.989 <0.01 22.917 (2) 6.121 0.206 − 0.983 <0.01 −26.498 (3) 0.063 0.986 19.78 0.953 <0.01 18.018 柠檬桉林 (1) 1.764 3.789 − 0.958 <0.01 47.513 (2) 4.427 0.302 − 0.997 <0.01 −38.749 (3) 0.102 0.985 25.82 0.975 <0.01 13.579 朴树林 (1) 2.873 1.713 − 0.990 <0.01 14.059 (2) 3.712 0.223 − 0.990 <0.01 −30.644 (3) 0.110 0.985 13.24 0.968 <0.01 15.682 ![]() 图 4 群落整体种-面积曲线最优拟合结果Figure 4. Best fitting results of species-area curve of the whole community
图 4 群落整体种-面积曲线最优拟合结果Figure 4. Best fitting results of species-area curve of the whole community![]() 图 5 灌木层种-面积曲线最优拟合结果Figure 5. Best fitting results of species-area curve of the shrub layer
图 5 灌木层种-面积曲线最优拟合结果Figure 5. Best fitting results of species-area curve of the shrub layer![]() 图 6 草本层种-面积曲线最优拟合结果Figure 6. Best fitting results of species-area curve of the herb layer
图 6 草本层种-面积曲线最优拟合结果Figure 6. Best fitting results of species-area curve of the herb layer从图4 ~ 图6的结果来看,随着样方的面积扩大,样方内的植物种类也随之增加,最初增长很快,以后逐渐缓慢,取样面积从300 ~ 400 m2物种数量增加量很小或几乎不再增加。图中的拟合曲线均比较靠近实践观测物种数量点,但图6中台湾相思群落灌木层种-面积拟合曲线与实际观测物种数量点拟合距离偏差较大,可能是因为琅岐岛台湾相思群落灌木层植物种类相对丰富。
2.3 基于最优模型的各群落3个对象物种多样性调查最小取样面积
选择拟合优度最好的幂函数的种-面积曲线拟合函数模型(2)所对应的最小取样面积方程(5)进行物种多样性调查的最小取样面积计算(表2)。因此,台湾相思林、柠檬桉林、朴树林的最小取样面积方程分别为:A台=(54.7ρ/11.813)1/0.269、A柠=(62.7ρ/10.642)1/0.307、A朴=(37.3ρ/7.774)1/0.271;灌木层的最小取样面积方程分别为:A台灌=(34.5ρ/5.804)1/0.314、A柠灌=(35.7ρ/6.069)1/0.309、A朴灌=(24.2ρ/3.992)1/0.309;草本层的最小取样面积方程分别为:A台草=(19.7ρ/6.121)1/0.206、A柠草=(25.7ρ/4.427)1/0.302、A朴草=(13.2ρ/3.712)1/0.223。
选择5种不同的估测精度计算3类海岛森林群落的取样面积。由表4可知,随着估测精度的增加,台湾相思林、桉树林、朴树林物种多样性调查的取样面积逐渐扩大,对应的种-面积曲线斜率逐渐减小。根据拐点法,在满足精度为60% ~ 90%的前提下,台湾相思林群落从精度70% ~ 80%时,曲线斜率由0.108变为0.09,即在精度为80%时,曲线斜率0.09 < 0.1,则台湾相思林调查的最小取样面积在79.2 ~ 130.1 m2;当取样精度为50%和60%时,此时的曲线斜率较大,且最小取样面积22.7和44.6 m2与实际明显不符。同理,台湾相思林灌木层、草本层的最小取样面积分别为57.4 ~ 93.7 m2(精度60% ~ 70%)和24.4 ~ 51.6 m2(精度60% ~ 70%);柠檬桉林群落及其灌木层、草本层的最小取样面积分别为100.6 ~ 155.6 m2(精度70% ~ 80%)、59.2 ~ 97.5 m2(精度60% ~ 70%)和34.1 ~ 62.3 m2(精度50% ~ 60%);朴树林群落及其灌木层、草本层的最小取样面积分别为49.5 ~ 87.4 m2(精度60% ~ 70%)、36.2 ~ 65.3 m2(精度50% ~ 60%)和13.2 ~ 29.9 m2(精度50% ~ 60%),可以将这些面积当作最小取样面积区间。
表 4 不同精度下琅岐岛3类森林群落调查的取样面积Table 4. Sample area of three forest communities in Langqi Island under different precision研究对象
Object of study森林
类型
Forest type不同估测精度 Different estimation precision ρ=0.5 ρ=0.6 ρ=0.7 ρ=0.8 ρ=0.9 取样面积
Sampling area / m²曲线斜率
Slope of curve取样面积
Sampling area / m2曲线斜率
Slope of curve取样面积
Sampling area / m2曲线斜率
Slope of curve取样面积
Sampling area / m2曲线斜率
Slope of curve取样面积
Sampling area / m2曲线斜率
Slope of curve群落整体 台湾相思林 22.7 0.324 44.6 0.198 79.2 0.130 130.1 0.090 201.5 0.066 柠檬
桉林33.8 0.285 61.1 0.189 101.0 0.133 156.1 0.099 229.0 0.076 朴树林 25.3 0.200 49.5 0.123 87.4 0.081 143.1 0.057 221 0.041 灌木层 台湾相思林 32.1 0.169 57.4 0.113 93.7 0.081 143.4 0.060 208.7 0.047 柠檬
桉林32.8 0.168 59.2 0.112 97.5 0.079 150.3 0.059 220 0.045 朴树林 36.2 0.103 65.3 0.069 107.5 0.049 165.6 0.036 242.5 0.028 草本层 台湾相思林 10.1 0.201 24.4 0.107 51.6 0.055 98.6 0.033 174.6 0.021 柠檬
桉林34.1 0.114 62.3 0.075 103.8 0.052 161.6 0.038 238.6 0.029 朴树林 13.2 0.111 29.9 0.059 59.7 0.035 108.7 0.022 184.3 0.014 当斜率为0.1时(具体拐点),台湾相思林群落及其灌木层、草本层的最小取样面积分别为113.5、68.9和24.4 m2,此时拟合曲线对应物种数分别为42.2、21.9和11.8,实际调查平均物种数分别为40.0 ~ 46.5、24.2 ~ 29.0和10.7 ~ 13.0;斜率为0.1时的柠檬桉林群落及其灌木层、草本层的最小取样面积分别为153.1、69.6和41.4 m2,此时拟合曲线对应物种数分别为49.9、22.5和13.6,实际调查平均物种数分别为49.3 ~ 58.8、21.7 ~ 28.8和13.2 ~ 15.5;斜率为0.1时的朴树林群落及其灌木层、草本层的最小取样面积分别为65.5、38和15.2 m2,此时拟合曲线对应物种数分别为24.1、12.3和6.8,实际调查平均物种数分别为24.3 ~ 30.3、12.0 ~ 14.5和6.0 ~ 7.2。可见,估测值与实际值均差别不大,表明曲线拟合程度较好,估测值与实际值均较为接近,因此可以将这些面积定义为最小取样面积(表5)。
表 5 斜率为0.1时的取样面积Table 5. Sample area with a slope of 0.1森林类型
Forest type斜率为0.1时的面积Area with a slope of 0.1 / m2 森林群落
Forest community灌木层
Shrub layer草本层
Herb layer台湾相思林 113.5 68.9 24.4 柠檬桉林 153.1 69.6 41.1 朴树林 65.5 38.0 15.2 3. 讨论
3.1 种-面积关系最优拟合模型分析
本研究设置了统一面积20 m × 20 m的调查样地来比较分析不同植物群落的种-面积关系和相应的最小取样面积。结果表明,对数函数模型、幂函数模型和逻辑斯蒂模型是比较适合巢式样方法的3种拟合曲线[5-9]。如果仅从R2大小判断拟合效果,台湾相思林草本层最优拟合模型为对数函数,朴树林草本层最优拟合模型为对数函数和幂函数。但从拟合优度检验的结果可知,琅岐岛3类海岛森林群落3个对象的赤池信息准则(AIC)最小值皆为幂函数模型,即3类森林群落3个对象的最优拟合模型都为幂函数。表明仅从R2的大小并不能准确判别拟合优度的好坏,还需要进一步的检验。最优模型为幂函数模型的原因可能是因为本研究选择样地面积都为20 m × 20 m(400 m2)的统一样地,以及在样地中特意选择了植被丰富、植物生长均匀的地方进行数据取样。同时许多学者已经关注到了种-面积曲线对样地尺度是具有依赖性的,如Williams[36]发现小尺度时的种-面积关系比较符合指数函数模型,中等尺度上的种-面积关系更符合幂函数模型,大尺度上时则更符合逻辑斯蒂模型[5, 6]。这很可能是因为样地面积(400 m2)属于中等尺度,才会得到幂函数为最优拟合模型的结果。姜俊等[37]和向悟生等[38]在分别对北温带针阔混交林的研究和在喀斯特和非喀斯特两种不同质地背景森林的研究也得到了相似的结论。本研究在对琅岐岛3类森林群落种-面积关系拟合的结果与Williams[36]和姜俊等[37]的研究结果一致,均表明在中等尺度时,幂函数模型的拟合效果最好。
3.2 琅岐岛3类森林群落最小取样面积分析
有研究利用不同方法来确定种-面积拟合后的最小取样面积。刘志兰等[24]以出现频率大于1的物种数占比70.4%作为取样精度来确定最小取样面积。法瑞学派常把包含总数80%的面积作为群落最小取样面积[4]。有研究认为“最小面积是能包括90%以上群落物种的面积”[39]。姜俊等[37]将种-面积曲线上斜率趋于稳定的那一点所对应的取样面积作为最小取样面积。杨利民等[40]将种-面积曲线上取样面积增加10%,而植物物种数增加不超过5%的拐点所对应的取样面积作为最小取样面积。赵鸿怡等[32]和任雪敏等[14]分别在研究香格里拉两种典型高寒草甸植物和太白山几种植物群落时,采用的不同研究精度计算最小取样面积及使用一定精度范围内的拐点法计算最小取样面积。本研究亦采用不同估测精度计算琅岐岛3类海岛森林群落3个对象的取样面积,能比较全面地提供不同研究需求下的最小取样面积选择。同时结合拐点法能够得到一定精度范围内较为确定的最小取样面积数值,方便人们参考。
群落最小取样面积是由乔、灌、草3个对象共同决定,但统计结果表明海岛乔木层种类较少,因此在相同取样精度下,群落灌木层、草本层很可能已经决定了群落物种数总体分布趋势。海岛3类森林群落调查的最小取样面积,随着估测精度的增加,取样面积也不断地增大,对应的种-面积曲线斜率逐渐减小。海岛3类森林群落(台湾相思林、柠檬桉林、朴树林)3个对象,在相同取样面积中的物种数量不同,在相同精度下的最小样地面积不相同,精度提高时所增加的取样面积变化幅度也不一样。从拐点法求得的最小取样面积结果也显示,3种森林群落的最小取样面积不尽相同。其他学者的研究也表明,相同样地面积不同群落类型的物种数量不同[41],不同群落所需的最小样地面积不同[12, 42]。因此在研究海岛森林群落生物多样性时,有必要将不同的群落类型及分成不同层次进行区分,从而准确地调查海岛森林群落植物多样性。
根据拐点法分析结果,在满足50% ~ 90%的精度下,平均物种总数(62.7)最多的柠檬桉林群落3个对象所需最小取样面积最大,平均物种总数(54.7)第2的台湾相思群落所需3个对象所需最小取样面积为第2,平均物种总数(37.3)最少的朴树林群落所需最小取样面积最小。可能与本研究在取样时选择植被生长均匀的样地有关,最小取样面积很可能随着物种数量的增多而增大,即最小取样面积跟群落物种数量存在一定的正相关性。而扬子松等[43]的研究发现,如果两个群落的物种丰富度相差不大,最小取样面积不一定随着物种丰富度的增大而增大,因为最小面积在受到物种丰富度影响的同时也会受到物种均匀度的影响。Keeley[44]认为不同物种丰度分布引起的群落组织结构差异可能导致不同的种-面积曲线,进而影响最小取样面积。相同精度的情况下,平均物种总数(62.5)最多的柠檬桉林群落,所需的最小样地面积最大,但平均物种总数(37.3)最少的朴树林群落比平均物种总数(54.7)第2的台湾相思群落在相同精度下所需的最小样地面积更大,灌木层和草本层也有类似的结果。可能是由于本研究设置了统一面积的调查样地,但由于台湾相思的固氮能力能够改善土壤环境,使得琅岐岛台湾相思林实际群落丰富度大于本研究设置样方范围的丰富度,导致台湾相思群落的最小取样面积比实际值偏小。且相比于台湾相思林样方,位于云龙村的朴树林样方受人为影响稍大,群落中偶见种的数量可能较多,而种-面积曲线只考虑物种种类的增加,会夸大偶见种地位,进而导致朴树群落最小取样面积增大。综上,最小取样面积会受各种因素的共同影响,如群落物种数量、群落差异、取样面积、人为干扰和偶见种等。
-
![]()
图 1 猕猴桃果园复合种植多花黄精
Figure 1. Polygonatum cyrtonema intercropping in an Actinidia chinensis orchard
![]()
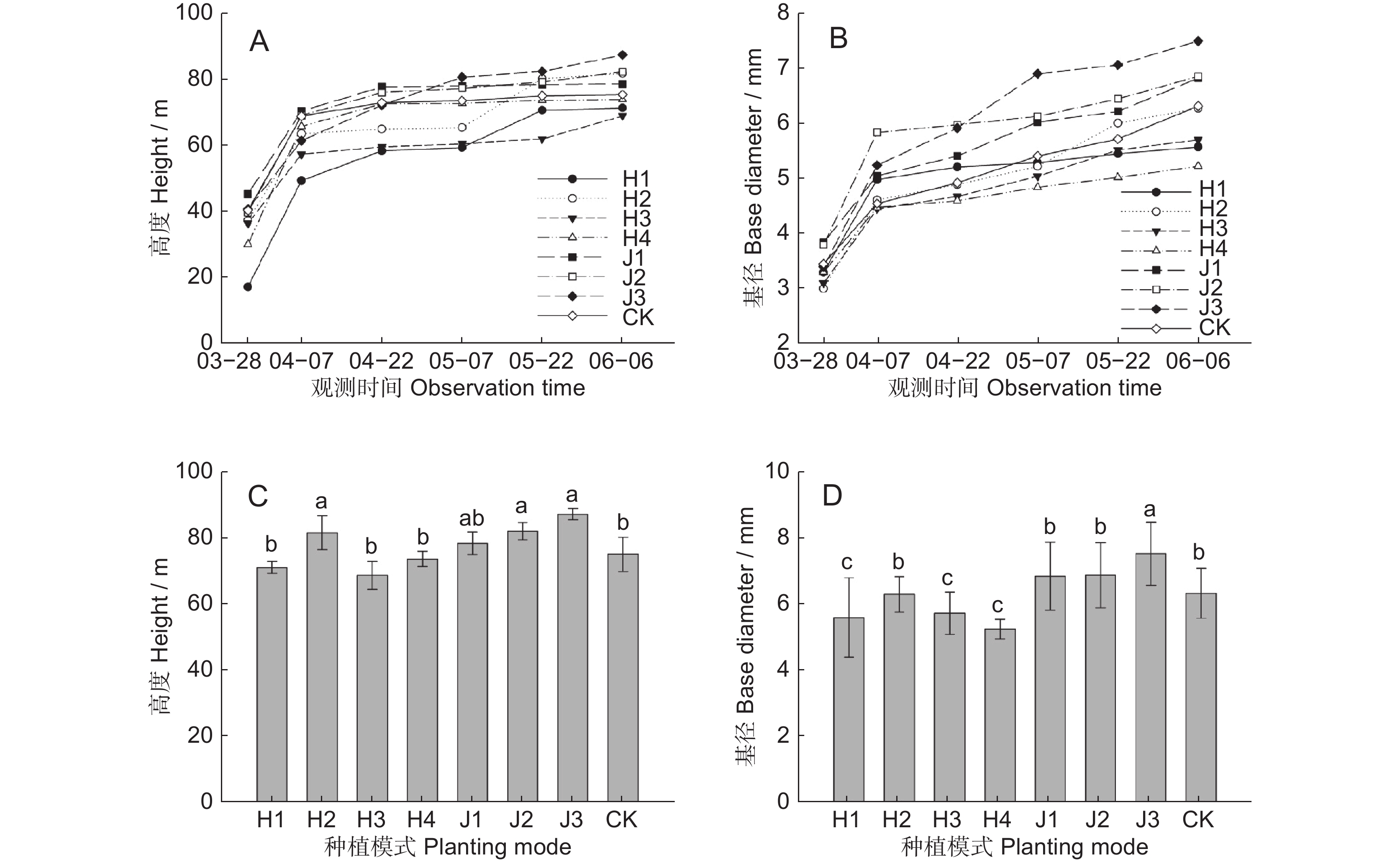
图 2 不同模式中多花黄精植株高度和基径生长变化
A:多花黄精生长期高度;B:生长期基径;C:6月上旬高度;D:6月上旬基径。不同小写字母表示差异显著(P<0.05),下同。
Figure 2. Changes in height and basal diameter of Polygonatum cyrtonema under different modes
A: Plant height during P. cyrtonema growth period; B: Basal diameter during growth period; C: Height in early June; D: Basal diameter in early June. Different lowercase letters indicate significant differences (P<0.05). Same below.
![]()
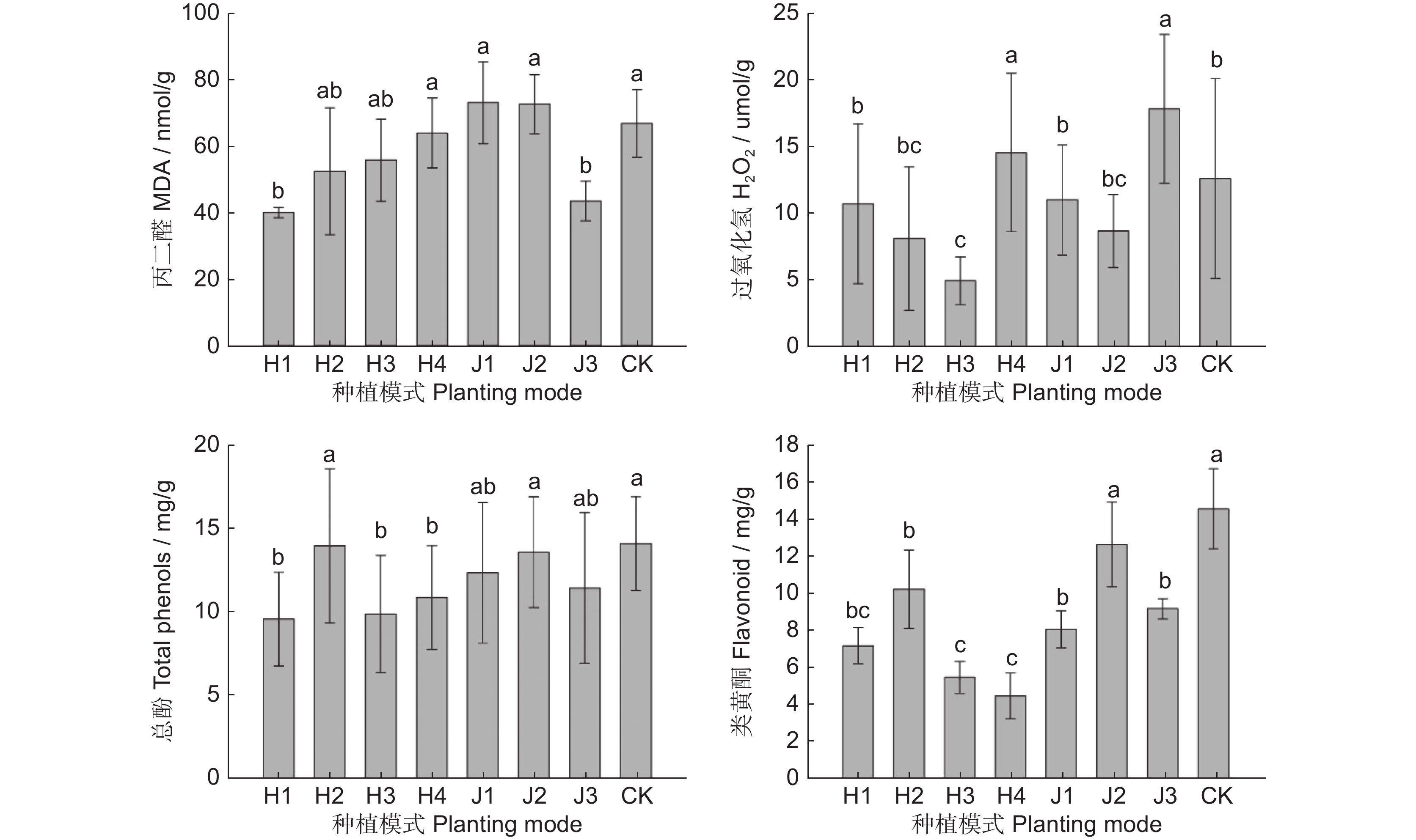
图 3 不同模式中多花黄精叶片丙二醛、过氧化氢及次生代谢物含量
Figure 3. Malondialdehyde, hydrogen peroxide, and secondary metabolite content in Polygonatum cyrtonema leaves in different modes
![]()
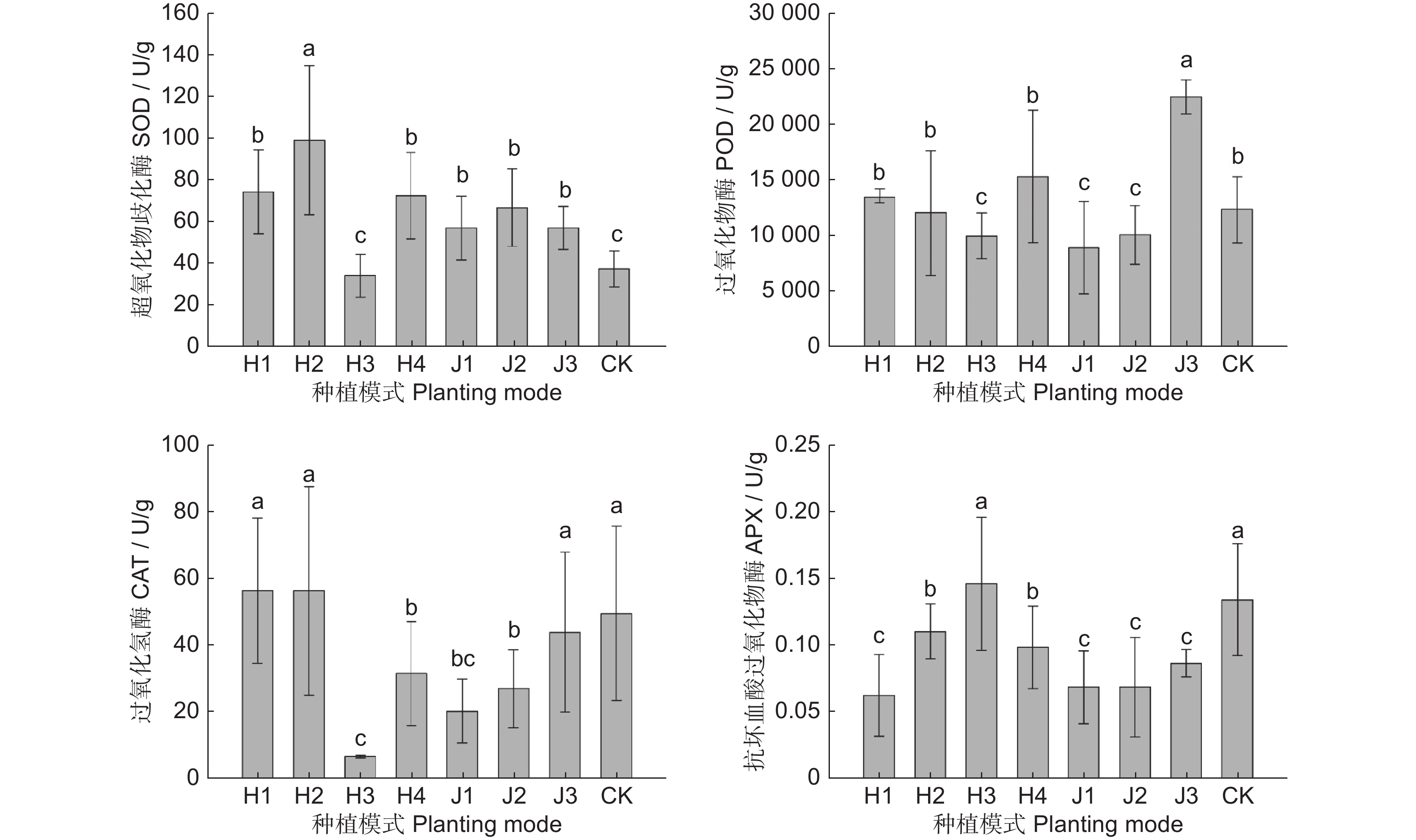
图 4 不同模式中多花黄精叶片抗氧化酶活性
Figure 4. Antioxidant enzyme activity in Polygonatum cyrtonema leaves in different modes
![]()
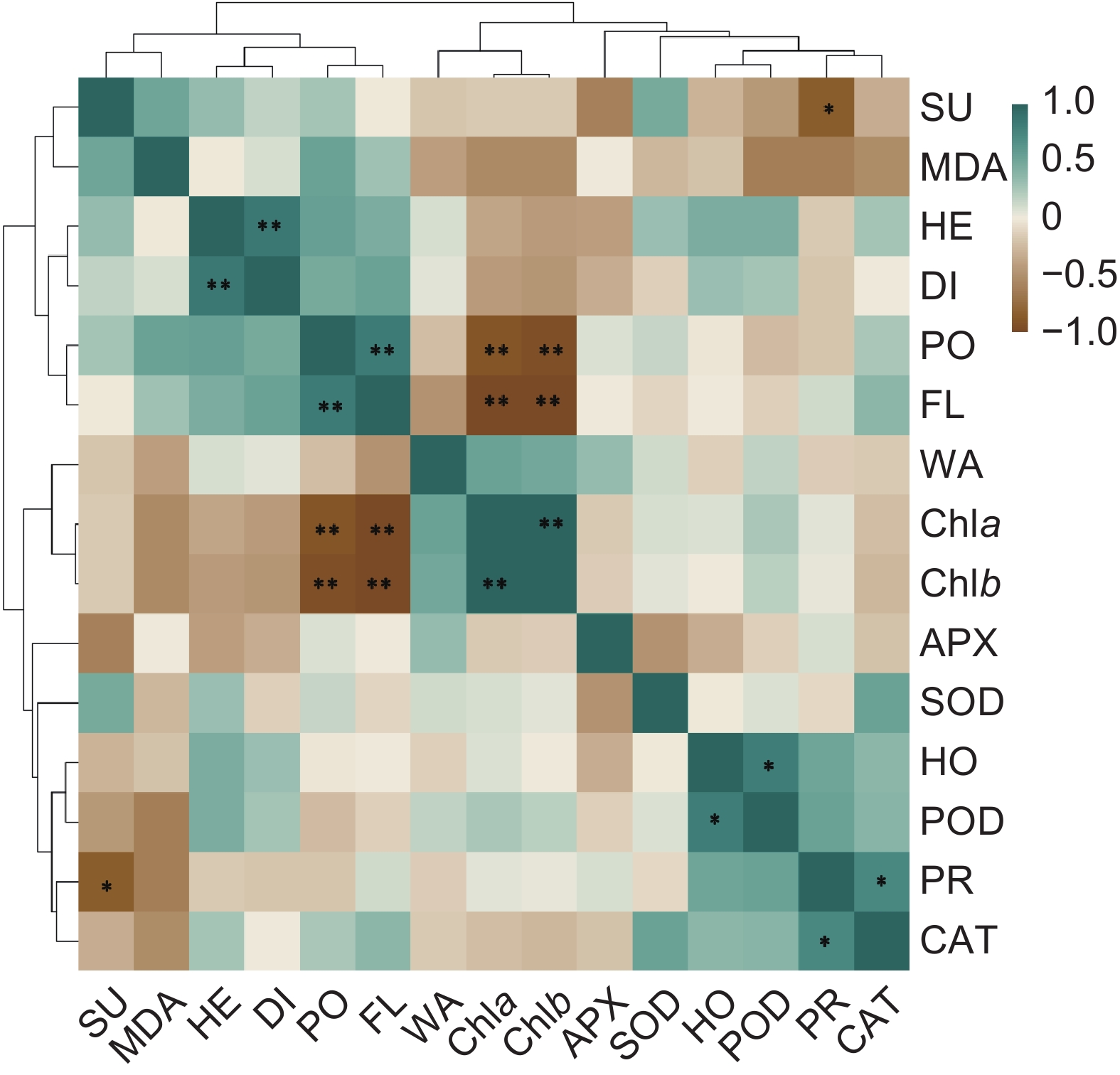
图 5 多花黄精生长生理指标相关性热图
SU:可溶性糖;MDA:丙二醛;HE:植株高度;DI:植株基径;PO:总酚;FL:类黄酮;WA:相对含水量;Chla:叶绿素a;Chlb:叶绿素b;APX:抗坏血酸过氧化物酶;SOD:超氧化物歧化酶;HO:过氧化氢;POD:过氧化物酶;PR:脯氨酸;CAT:过氧化氢酶。*:P<0.05;**:P<0.01。
Figure 5. Correlation heatmap of Polygonatum cyrtonema growth and physiological indices
SU: Soluble sugar; MDA: Malondialdehyde; HE: Height; DI: Basal diameter; PO: Total phenols; FL: Flavonoid; WA: Relative water content; Chla: Chlorophyll a; Chlb: Chlorophyll b; APX: Ascorbate peroxidase; SOD: Superoxide dismutase; HO: H2O2; POD: Peroxidase; PR: Proline; CAT: Catalase.
![]()
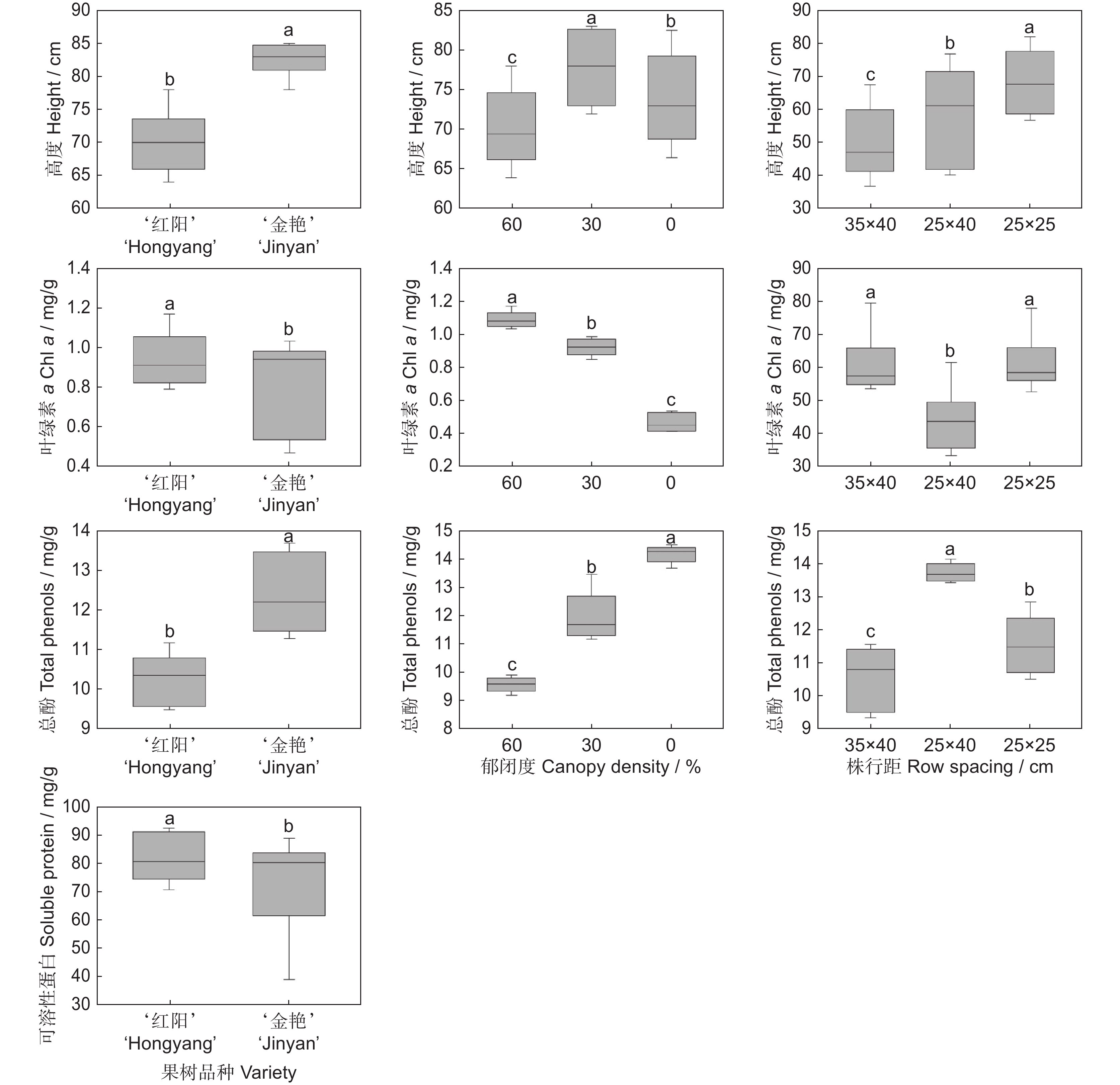
图 6 猕猴桃品种、郁闭度及种植密度对多花黄精生长生理适应性的影响
Figure 6. Effects of fruit variety, canopy density, and planting density on Polygonatum cyrtonema growth and physiological adaptability
表 1 猕猴桃-多花黄精复合种植模式设置
Table 1 Setup of Actinidia chinensis and Polygonatum cyrtonema intercropping system
编号
Number果树品种
Variety郁闭度
Canopy density / %光照强度
Light Intensity / lux光量子通量密度
Photon flux density / µmol·m2·s株行距
Row spacing / cmH1 ‘红阳’ 60 30 875±3 450 702±307 25×40(中密度) H2 ‘红阳’ 30 60 788±3 319 1 133±305 25×40(中密度) H3 ‘红阳’ 30 35×40(低密度) H4 ‘红阳’ 30 25×25(高密度) J1 ‘金艳’ 30 59 246±2 136 1 111±288 25×25(高密度) J2 ‘金艳’ 30 25×40(中密度) J3 ‘金艳’ 30 35×40(低密度) CK − 0 84 206±5 032 1 470±330 25×40(中密度) 注:‘金艳’盛果期果园树势较强,郁闭度一般在70%以上,林下光照条件不适宜多花黄精生长,因而只选择了幼龄果园作为研究对象。 Notes: During the peak fruit-bearing period, trees in the ‘Jinyan’ orchard exhibited strong growth, with a canopy density generally exceeding 70%. Understory light conditions were unsuitable for P. cyrtonema growth; therefore, only young orchards were selected for this study.  下载: 导出CSV
下载: 导出CSV
表 2 不同模式中多花黄精叶片含水率、叶绿素及渗透调剂物质含量
Table 2 Water, chlorophyll, and osmotic compound content in Polygonatum cyrtonema leaves in different modes
模式
Mode相对含水量
Relative water content / %叶绿素a
Chlorophyll a / mg/g叶绿素b
Chlorophyll b / mg/g可溶性糖
Soluble sugar / mg/g可溶性蛋白
Soluble protein / mg/g脯氨酸
Proline / μg/gH1 85.27±1.78b 1.09±0.07a 0.54±0.04a 26.02±6.52c 96.55±9.69a 267.00±69.80b H2 91.30±2.30a 0.79±0.03ab 0.38±0.02b 41.15±7.80b 80.75±23.01b 345.50±19.78a H3 90.54±5.69a 1.06±0.29a 0.53±0.20a 23.15±5.36c 75.05±4.48b 279.61±41.34b H4 87.01±4.74ab 1.08±0.25a 0.52±0.14a 38.16±3.28b 82.74±9.36b 284.05±20.30b J1 88.64±5.70ab 0.96±0.02a 0.46±0.02a 48.80±9.32b 73.38±15.94b 276.70±49.31b J2 82.99±6.92b 0.52±0.05b 0.26±0.02c 61.91±14.11a 67.53±24.85b 231.40±21.63c J3 89.87±3.48a 0.97±0.07a 0.46±0.06a 22.99±6.23c 90.36±14.72ab 246.62±25.63c CK 84.36±5.06b 0.46±0.07b 0.23±0.03c 12.07±3.32d 96.63±10.74a 315.93±42.11a 注:同列不同小写字母表示差异显著,P<0.05。 Note: Different lowercase letters in the same column indicate significant differences, P<0.05.
下载: 导出CSV
表 3 多花黄精生长生理指标主成分分析
Table 3 Principal component analysis of Polygonatum cyrtonema growth and physiological indices
指标 Index PC1 PC2 PC3 方差百分比 / % 33.256 24.301 17.026 高度 0.559 0.502 0.548 基径 0.580 0.343 0.337 相对含水量 −0.459 0.023 0.227 叶绿素a −0.934 −0.008 0.328 叶绿素b −0.952 −0.063 0.283 可溶性糖 0.398 −0.464 0.761 可溶性蛋白 −0.252 0.766 −0.510 H2O2 −0.010 0.771 0.147 MDA 0.603 −0.613 −0.003 总酚 0.923 0.064 −0.041 类黄酮 0.901 0.249 −0.294 SOD 0.035 0.229 0.620 APX −0.108 −0.191 −0.735 POD −0.251 0.850 0.139 CAT 0.151 0.789 −0.089
下载: 导出CSV
表 4 不同模式对多花黄精生长生理适应性影响的多因素方差分析
Table 4 Multivariate analysis of variance on effects of different modes on Polygonatum cyrtonema growth and physiological adaptability
组合因子
Factors高度
Height叶绿素a
Chlorophyll a可溶性蛋白
Soluble protein丙二醛
MDA总酚
Total phenols过氧化物酶
POD果树品种 0.003** 0.030* 0.010* 0.303 0.010* 0.499 果园郁闭度 0.041* 0.022* 0.745 0.178 0.008** 0.707 多花黄精种植密度 0.132 0.005** 0.231 0.026* 0.005** 0.115 品种×郁闭度 − − 0.642 − − − 品种×密度 0.054 0.491 − 0.055 0.087 0.426 郁闭度×密度 − − 0.256 − − − 品种×郁闭度×密度 − − − − − − Notes: *, P<0.05; **, P<0.01.
下载: 导出CSV
-
[1] 罗敏,章文伟,邓才富,谭秋生,罗川,罗舜. 药用植物多花黄精研究进展[J]. 时珍国医国药,2016,27(6):1467−1469. Luo M,Zhang WW,Deng CF,Tan QS,Luo C,Luo S. Advances in studies of medicinal crop Polygonatum cyrtonema Hua[J]. Lishizhen Medicine and Materia Medica Research,2016,27(6):1467−1469.
[2] 陈炬烽,钟起尧,郭志鹏,林燕华. 人工生态种植对土壤理化性质、多花黄精产量及品质的影响[J]. 中药材,2023,46(7):1613−1616. [3] 万修福,杨野,康传志,王升,王铁霖,等. 林草中药材生态种植现状分析及展望[J]. 中国现代中药,2021,23(8):1311−1318. Wan XF,Yang Y,Kang CZ,Wang S,Wang TL,et al. Current situation and future perspectives of ecological planting of Chinese medicinal plants in forests and grasslands[J]. Mod Chin Med,2021,23(8):1311−1318.
[4] Brooker RW,Bennett AE,Cong WF,Daniell TJ,George TS,et al. Improving intercropping:a synthesis of research in agronomy,plant physiology and ecology[J]. New Phytol,2015,206(1):107−117. doi: 10.1111/nph.13132
[5] 刘跃钧,蒋燕锋,葛永金,姚理武,谢建秋,等. 锥栗-多花黄精不同复合经营模式经济生态效益评价[J]. 经济林研究,2020,38(4):72−81. Liu YJ,Jiang YF,Ge YJ,Yao LW,Xie JQ,et al. Economic and ecological benefits evaluation of different compound management modes of Castanea henryi and Polygonatum cyrtonema[J]. Non-Wood Forest Research,2020,38(4):72−81.
[6] 黄云鹏,范繁荣,王邦富,沈琼桃,陈绍煌,等. 4种不同林分类型对多花黄精生长的影响[J]. 西部林业科学,2016,45(5):132−135. Huang YP,Fan FR,Wang BF,Shen QT,Chen SH,et al. The influence of four different forest types on the growth of Polygonatum cyrtonema Hua[J]. Journal of West China Forestry Science,2016,45(5):132−135.
[7] 卢玉鹏,高柱,张小丽,陈璐,王小玲. 果园生态系统复合经营的开展模式及生态机制综述[J]. 中国果树,2021(12):9−15. Lu YP,Gao Z,Zhang XL,Chen L,Wang XL. Review on the development model and ecological mechanism of compound management of orchard ecosystem[J]. China Fruits,2021(12):9−15.
[8] Wang YJ,Liu L,Luo Y,Awasthi MK,Yang JF,et al. Mulching practices alter the bacterial-fungal community and network in favor of soil quality in a semiarid orchard system[J]. Sci Total Environ,2020,725:138527. doi: 10.1016/j.scitotenv.2020.138527
[9] Gómez-Muñoz B,Hatch DJ,Bol R,García-Ruiz R. Nutrient dynamics during decomposition of the residues from a sown legume or ruderal plant cover in an olive oil orchard[J]. Agric Ecosyst Environ,2014,184:115−123. doi: 10.1016/j.agee.2013.11.020
[10] 秦秦,宋科,孙丽娟,孙雅菲,王峻,等. 猕猴桃园行间生草对土壤养分的影响及有效性评价[J]. 果树学报,2020,37(1):68−76. Qin Q,Song K,Sun LJ,Sun YF,Wang J,et al. Effect of inter-row sod system on the contents and availability of soil nutrients in a kiwifruit orchard[J]. Journal of Fruit Science,2020,37(1):68−76.
[11] Michel L,Peña Á,Pastenes C,Berríos P,Rombolà AD,et al. Sustainable strategies to prevent iron deficiency,improve yield and berry composition in blueberry (Vaccinium spp.)[J]. Front Plant Sci,2019,10:255. doi: 10.3389/fpls.2019.00255
[12] 赵俊侠,张中社,龙凤来. 猕猴桃果园套种板蓝根试验研究[J]. 价值工程,2014,33(24):315−316. Zhao JX,Zhang ZS,Long FL. A testing study on the interplanting of Radix isatidis in the kiwifruit orchards[J]. Value Engineering,2014,33(24):315−316.
[13] 张彩霞. 天水地区樱桃——药材果园复合生态农业模式及其效益分析[J]. 现代农业科技,2015(4):114−115. doi: 10.3969/j.issn.1007-5739.2015.04.070 [14] 高秋美,任丽华,米真如,孟庆峰,董秋颖,等. 不同光照强度对多花黄精生长及光合特性的影响[J]. 山东农业科学,2021,53(6):44−47. Gao QM,Ren LH,Mi ZR,Meng QF,Dong QY,et al. Effects of different illumination intensity on growth and photosynthetic characteristics of Polygonatum cyrtonema Hua[J]. Shandong Agricultural Sciences,2021,53(6):44−47.
[15] 郭妮. 栽培措施对林下多花黄精产量和品质的影响[D]. 重庆:西南大学,2019:40−42. [16] 倪松娟. 毛竹林不同生境下多花黄精生长及其与土壤性质的关系[D]. 南昌:江西农业大学,2021:14−15. [17] Lu YP,Gao Z,Mao JP,Chen L,Zhang XL,et al. Litter decomposition characteristics and variety differences in a kiwifruit orchard in subtropical climate zone of China[J]. Agronomy,2023,13(3):774. doi: 10.3390/agronomy13030774
[18] Gao SS,Wang YL,Yu S,Huang YQ,Liu HC,et al. Effects of drought stress on growth,physiology and secondary metabolites of two Adonis species in Northeast China[J]. Sci Hortic,2020,259:108795. doi: 10.1016/j.scienta.2019.108795
[19] 张维,付复华,罗赛男,赖灯妮,朱向荣,张群. 湖南红心猕猴桃品种品质评价及综合分析[J]. 食品与发酵工业,2021,47(5):201−210. Zhang W,Fu FH,Luo SN,Lai DN,Zhu XR,Zhang Q. Quality analysis and evaluation of Hunan red kiwifruit varieties[J]. Food and Fermentation Industries,2021,47(5):201−210.
[20] 赵海云,姜成英,戚建莉,赵梦炯,金高明,吴文俊. 油橄榄落果规律及果实发育期叶片可溶性糖、淀粉、内源激素含量变化[J]. 中国果树,2022(10):43−48. Zhao HY,Jiang CY,Qi JL,Zhao MJ,Jin GM,Wu WJ. The law of fruit dropping and changes of soluble sugar,starch and endogenous hormone contents in leaves of olive during fruit development period[J]. China Fruits,2022(10):43−48.
[21] 韩富亮,袁春龙,郭安鹊,张予林,李运奎. 二喹啉甲酸法(BCA)分析蛋白多肽的原理、影响因素和优点[J]. 食品与发酵工业,2014,40(11):202−207. Han FL,Yuan CL,Guo AQ,Zhang YL,Li YK. The principle,influence factors and advantages of bicinchoninic acid method (BCA) for protein and peptide assay[J]. Food and Fermentation Industries,2014,40(11):202−207.
[22] 王小玲,于一尊,高柱,黄国昌,刘小宁. 羽扇豆套种对猕猴桃叶片和果实丙二醛含量的影响[J]. 北方园艺,2018(24):54−58. Wang XL,Yu YZ,Gao Z,Huang GC,Liu XN. Effects of intercropping Lupin on malondialdehyde content in kiwifruit leaf and fruit[J]. Northern Horticulture,2018(24):54−58.
[23] 郑飞雪,张新忠,王忆,韩振海. 苹果实生树个体发育不同阶段叶片H2O2含量及相关酶活性的变化[J]. 果树学报,2013,30(5):759−764. Zheng FX,Zhang XZ,Wang Y,Han ZH. Changes of H2O2 content and activities of related enzymes in leaf among ontogenetic phases in apple (Malus ssp. )[J]. Journal of Fruit Science,2013,30(5):759−764.
[24] 郝岩,王英平,李金玲,金银萍. 不同采收时期五味子叶片中总黄酮、总黄酮醇和总酚酸含量的动态变化研究[J]. 时珍国医国药,2021,32(10):2500−2502. doi: 10.3969/j.issn.1008-0805.2021.10.52 [25] Caverzan A,Passaia G,Rosa SB,Ribeiro CW,Lazzarotto F,et al. Plant responses to stresses:role of ascorbate peroxidase in the antioxidant protection[J]. Genet Mol Biol,2012,35(4 Suppl):1011−1019.
[26] Zhang ZY,Liu HH,Sun C,Ma QB,Bu HY,et al. A C2H2 zinc-finger protein OsZFP213 interacts with OsMAPK3 to enhance salt tolerance in rice[J]. Journal of Plant Physiology,2018,229:100−110. doi: 10.1016/j.jplph.2018.07.003
[27] 王艺,韦小丽. 不同光照对植物生长、生理生化和形态结构影响的研究进展[J]. 山地农业生物学报,2010,29(4):353−359,370. Wang Y,Wei XL. Advance on the effects of different light environments on growth,physiological biochemistry and morphostructure of plant[J]. Journal of Mountain Agriculture and Biology,2010,29(4):353−359,370.
[28] 王梅,徐正茹,张建旗,曹效东,刘乐乐,等. 遮阴对10种野生观赏植物生长及生理特性的影响[J]. 草业科学,2017,34(5):1008−1016. Wang M,Xu ZR,Zhang JQ,Cao XD,Liu LL,et al. Effect of shades on growth and physiological characteristics of 10 species of wild ornamental plants in Lanzhou[J]. Pratacultural Science,2017,34(5):1008−1016.
[29] 童龙,张磊,高勇军,陈丽洁,耿养会,李彬. 不同遮阴处理对多花黄精生理生长的影响[J]. 西南林业大学学报,2020,40(3):68−75. Tong L,Zhang L,Gao YJ,Chen LJ,Geng YH,Li B. Growth and physiological characteristics of Polygonatum cyrtonema under different shading treatments[J]. Journal of Southwest Forestry University,2020,40(3):68−75.
[30] Liu SL,Yang RJ,Ren B,Wang MH,Ma MD. Differences in photosynthetic capacity,chlorophyll fluorescence,and antioxidant system between invasive Alnus formosana and its native congener in response to different irradiance levels[J]. Botany,2016,94(12):1087−1101. doi: 10.1139/cjb-2016-0026
[31] 何小燕,马锦林,张日清,叶航. 弱光胁迫对植物生长影响的研究进展[J]. 经济林研究,2011,29(4):131−136. He XY,Ma JL,Zhang RQ,Ye H. Research progress on effects of low-light stress on plant growth[J]. Non-Wood Forest Research,2011,29(4):131−136.
[32] 丁凯鑫,王立春,田国奎,王海艳,李凤云,等. 外源烯效唑对干旱胁迫下马铃薯叶片抗氧化能力及渗透调节的影响[J]. 核农学报,2024,38(1):169−178. doi: 10.11869/j.issn.1000-8551.2024.01.0169 Ding KX,Wang LC,Tian GK,Wang HY,Li FY,et al. Effect of exogenous uniconazole on antioxidant capacity and osmotic adjustment of potato leaves under drought stress[J]. Journal of Nuclear Agricultural Sciences,2024,38(1):169−178. doi: 10.11869/j.issn.1000-8551.2024.01.0169
[33] Cheng L,Han M,Yang LM,Li Y,Sun Z,et al. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress[J]. Industrial Crops and Products,2018,122:473−482. doi: 10.1016/j.indcrop.2018.06.030
[34] Zhu DB,Hu KD,Guo XK,Liu Y,Hu LY,et al. Sulfur Dioxide Enhances Endogenous Hydrogen sulfide accumulation and alleviates oxidative stress induced by aluminum stress in germinating wheat seeds[J]. Oxid Med Cell Longev,2015,2015:612363.
[35] 蔡元保,杨祥燕,孙光明,张治礼,黄强,等. 菠萝叶片色泽、色素及抗氧化活性的关系[J]. 植物科学学报,2017,35(2):283−290. Cai YB,Yang XY,Sun GM,Zhang ZL,Huang Q,et al. Relationship among colors,pigments,and antioxidant activities of pineapple leaves[J]. Plant Science Journal,2017,35(2):283−290.
[36] 卢发光,顾立峰,刘昱茜,任桢,施雨,等. 种植密度和施氮量互作对盐碱地紫花苜蓿生长性能和生理特性的影响[J]. 草业科学,2021,38(8):1570−1578. Lu FG,Gu LF,Liu YQ,Ren Z,Shi Y,et al. Interaction effects of plant density and nitrogen rate on the growth and physiological traits of alfalfa in saline alkali soils[J]. Pratacultural Science,2021,38(8):1570−1578.
[37] 李秀亚,宋福兵,肖钧,汪志威,韩振诚. 5个猕猴桃品种在六盘水的生物学特性比较[J]. 安徽农业科学,2019,47(17):47−51. doi: 10.3969/j.issn.0517-6611.2019.17.014 Li XY,Song FB,Xiao J,Wang ZW,Han ZC. Comparison of biological characteristics of five kiwi varieties in Liupanshui[J]. Journal of Anhui Agricultural Sciences,2019,47(17):47−51. doi: 10.3969/j.issn.0517-6611.2019.17.014
[38] 卢瑜珺. 阿拉尔骏枣和灰枣根系年生长变化与地上部生长结果的关系研究[D]. 阿拉尔:塔里木大学,2022:12−32. [39] 张春兰,李苇洁,姚红艳,罗充,徐旵旵,申笑阳. 不同猕猴桃品种根际AM真菌多样性与土壤养分相关性分析[J]. 果树学报,2017,34(3):344−353. Zhang CL,Li WJ,Yao HY,Luo C,Xu HH,Shen XY. Correlation study on the diversity of the AM fungi and soil nutrients in the rhizosphere of different kiwifruit cultivars[J]. Journal of Fruit Science,2017,34(3):344−353.







计量
- 文章访问数: 124
- HTML全文浏览量: 25
- PDF下载量: 9
