
Age structure and demographic dynamics of Ammopiptanthus nanus (M. Pop.) Cheng f. populations
-
摘要:
新疆沙冬青(Ammopiptanthus nanus (M. Pop.) Cheng f.)为第三纪古地中海沿岸的珍稀孑遗物种,是国家Ⅱ级保护植物。本研究通过种群年龄结构、静态生命表、存活曲线、生存函数曲线及时间序列模型分析,揭示新疆沙冬青种群年龄结构及数量动态趋势。结果显示,新疆沙冬青种群老年个体数量最多,占总株数的48.1%,幼苗占比最低,仅为7.0%,种群年龄结构呈衰退型;存活曲线呈Deevey-Ⅱ型,种群总体死亡率较稳定;种群数量具有前期锐减、中期稳定、后期衰退的动态趋势,表明新疆沙冬青有较高的灭绝风险。增加种群幼苗数量,提高幼龄级到成年龄级过渡阶段的存活率,是促进新疆沙冬青良性发展的关键措施。
Abstract:Ammopiptanthus nanus (M. Pop.) Cheng f. is a rare Tertiary relict species that originated along the Tethys coast and is currently classified as a state Grade Ⅱ protected plant due to demographic scarcity. This study examined the population age structure and demographic dynamics through analyses of static life tables, survival curves, survival function curves, and time series models. Results showed that mature individuals constituted the largest proportion of the population (48.1%), while saplings accounted for the lowest proportion (7.0%), reflecting a declining age structure. The species exhibited a Deevey-Ⅱ survival curve, with a stable overall mortality rate. However, a pronounced decline in the early stages, followed by a stabilization phase in the middle period and a subsequent decline in the later stages, suggests a heightened risk of extinction. Enhancing seedling recruitment and improving the survival rate during the transition from juvenile to adult stages are critical for ensuring population sustainability and long-term viability.
-
Keywords:
- Ammopiptanthus nanus /
- Survival analysis /
- Time series models /
- Population dynamics
-
青藏高原被誉为“世界屋脊”,这里有着极其复杂的地理环境、多样的生态系统和独特的生物多样性资源[1],同时也是对全球变暖最敏感的地区之一[2]。自20世纪中期以来,人类活动和气候环境变化对青藏高原的影响日益加剧[3-6],全球变暖导致的温度上升和降水模式的改变,可能对青藏高原生物多样性的组成和分布产生显著影响[7, 8]。垫状植物是高寒生态系统中的关键物种和先锋物种[9],对气候变化特别敏感[10],当前的气候变化可能会威胁到高山垫状植物的生存[11]。垫状植物具有一定的保温作用,这种特性被认为是其能够适应高海拔极端环境的重要因素。Körner和Demoraes[12]利用红外温度计,在晴天对垫状植物Silence acaulis L. 进行了12 h的温度记录,发现其表面温度远远高于气温。Fisher和Kuhn[13]在夏季的晴天,用热敏电阻器温度计插入不同深度,记录垫状半灌木植物 Diapensia lapponica L.的表面和内部温度,结果发现在一昼夜中其表面温度比气温变化的更快,内部温度变化则较平缓。还有研究表明,垫状植物能改善其基部土壤的温度和湿度,提高其他草本植物幼苗的存活率[14],并能有效降低自身周围的风速,减少对流带来的热损失[15]。但先前研究的垫状植物大多为高山藓类植物,且未说明决定垫状植物自身保温作用的关键因素,垫状植物保温作用的相关研究还有待深入,这也是认识和理解其先锋作用和适应性进化的基础。
垫状点地梅(Androsace tapete Maxim.)是青藏高原最典型的垫状植物,具有密集的带小叶子的莲座组成紧密的冠层,贴近地面紧凑的半球形垫状结构,能够减少风的机械应力,缓冲温度波动[16]。垫状点地梅也是分布海拔最高的种子植物,生长在高寒、干旱、昼夜温差大的恶劣环境中,其最高覆盖率的海拔为4900 ~ 4950 m,最适生长温度为9℃ ~ 14℃,在生长季能够耐受−10℃ ~ −5℃的季节性霜冻[17]。垫状点地梅具有可以在较温暖年份提前长出叶片等待季风降雨到来的物候策略,从而避免季风前干旱和霜冻造成的损害[18]。在其垫层内部能够营造一个温度比周围空气更稳定的微环境[19],为其他植物提供生长基质并减少水分损失,促进它们的生长和定殖[20],被誉为“高原生态系统工程师”。因此,垫状点地梅对极端温度的适应性备受学者的关注。
关于垫状点地梅的保温作用,许多研究均表明垫状植物的温度高于气温[12, 13],认为垫状体结构具有保温作用。而闫巍[21]则认为垫状点地梅是一个被动吸热器,其本身并没有所谓的“保温功能”。由于各研究使用的测温仪器、研究物种、测温时长及测温部位等的不同,多以垫状植物表面温度作为整个垫状体的温度。例如闫巍[21]以垫状点地梅表面下0.5 cm处的温度作为垫状点地梅温度,且气温和地表温度数据来源于同一海拔高度的简易气象站资料,并非同时于样本附近测量,因此,不同研究得到的结论存在争议。目前,对垫状点地梅的研究主要集中在区系类型和分布特征[22-24]、形态特征和生态适应性[25-27]以及空间遗传结构[28, 29]等方面。关于垫状点地梅的保温作用及其影响因素仍有待进一步探索。
垫状植物的面积越大,其温度变化幅度就越大[21],其保温能力可能与垫状结构的紧密程度及其大小有关,但不排除其自身代谢对温度的影响。实地采样中发现,部分垫状点地梅居群中存在一些死亡个体,这些个体已失去生理活性,但保留了完整的垫状结构。因此,通过测定垫状点地梅存活与死亡个体的内部温度,并进行比较,可能揭示影响其保温作用的主要因素。在青藏高原高海拔地区,植物的生长季可能会出现极端的低温天气,如突发的冰雹等。未经冷驯化的垫状点地梅如何在抗冻能力最弱时抵御温度的剧烈变化?其垫状结构是怎样发挥保温作用的? 经冷驯化后,冬天垫状点地梅的保温能力是否增强? 为了探讨这些问题,本研究测量了夏季和冬季垫状点地梅的表面温度和内部温度、附近气温以及土壤温度的昼夜变化,分析了低温处理对垫状点地梅表面及内部温度的影响,旨在阐明垫状点地梅对极端温度变化的适应性。
1. 材料与方法
1.1 实验材料
选取西藏不同地区的垫状点地梅作为实验材料,样点信息、所测项目和测温时长见表1。用热敏电阻型温度计(WS-C-PDP,分辨率为0.01℃,误差为0.1℃)测定多组直径相似、结构完整、位置相近的垫状点地梅存活与死亡个体的内部温度(图1)。用热敏电阻型自动温度记录仪(CHL-TF20,分辨率为0.01℃,误差为0.1℃,采集频率为1次/ min)。在夏季7月沿500 m左右的海拔梯度连续监测夺底沟、羊八井和麦地卡的垫状点地梅居群的温度数据,在冬季11月连续监测米拉山的垫状点地梅存活与死亡个体的温度数据。
表 1 实验样点信息及所测项目Table 1. Information on experimental sites and measured items样点
Site经纬度
Latitude and longitude海拔
Altitude / m测温月份
Month of measurement测温项目
Temperature measurement item拉孜热萨乡 29.072274°N
87.962344°E4408 7月 垫状点地梅存活和死亡个体的内部温度(共25组) 聂拉木门布乡 28.657388°N
86.123398°E4598 7月 垫状点地梅存活和死亡个体内部温度、
土壤5 cm处温度(共28组)拉萨夺底沟 29.716398°N
91.180987°E3898 7月 垫状点地梅表面温度、内部温度、附近气温和
土壤5 cm处温度的昼夜变化当雄羊八井 30.072869°N
90.433037°E4404 7月 垫状点地梅表面温度、内部温度、附近气温和
土壤5 cm处温度的昼夜变化那曲麦地卡湿地 31.180789°N
93.014379°E4947 7月 垫状点地梅表面温度、内部温度、附近气温和
土壤5 cm处温度的昼夜变化拉萨米拉山 29.798365°N
92.341356°E4808 11月 垫状点地梅存活与死亡个体的表面温度、内部温度、附近气温和土壤5 cm处温度的昼夜变化 ![]() 图 1 青藏高原垫状点地梅测温示意图A:垫状点地梅死亡个体内部温度的测量;B:垫状点地梅存活个体内部温度的测量;C:垫状点地梅附近土温的测量。Figure 1. Temperature measurement of Androsace tapete on Qinghai-Tibet PlateauA: Internal temperature measurement of dead A. tapete individuals; B: Internal temperature measurement of living A. tapete individuals; C: Soil temperature measurement near A. tapete.
图 1 青藏高原垫状点地梅测温示意图A:垫状点地梅死亡个体内部温度的测量;B:垫状点地梅存活个体内部温度的测量;C:垫状点地梅附近土温的测量。Figure 1. Temperature measurement of Androsace tapete on Qinghai-Tibet PlateauA: Internal temperature measurement of dead A. tapete individuals; B: Internal temperature measurement of living A. tapete individuals; C: Soil temperature measurement near A. tapete.连续测温的项目包括垫状点地梅表面和内部温度,以及其附近的气温和土壤温度。所选择测温的垫状点地梅个体直径均为20 cm左右,气温的测定是将温度记录仪探头放置于垫状点地梅附近距地面约20 cm的空气中(日间受太阳辐射的影响,其温度数值会略高于气温实际值,夜间的气温数据更为准确),表面温度的测温探头斜插入垫状点地梅表面下约0.5 cm处,测量内部温度和土壤温度的探头从表面插入5 cm,每个观测点各测3个个体。
此外,为模拟青藏高原突发的冰雹天气,在夏季的晴天对垫状点地梅以及附近的土壤同时撒下碎冰,做3次低温处理,撒冰量约0.3 × 10−3 m3,用温度记录仪记录整个过程中垫状点地梅表面、内部温度和土壤温度的动态变化。为探究垫状点地梅的保温能力是否受海拔变化、自身生理活性和冷驯化的影响,将不同海拔、不同生存状态、不同季节的垫状点地梅保温能力进行比较。通过垫状点地梅附近气温与其内部温度最大变化幅度的比值来表示其保温能力,HC =
ATmax−ATminITmax−ITmin ,其中HC为保温能力,当HC > 1时,认为该物体具有保温能力,其中ATmax为地表20 cm处所测的最高气温,ATmin为所测的最低气温,ITmax为垫状点地梅内部5 cm处所测的最高温度,ITmin为所测的内部最低温度。1.2 数据处理
数据处理和分析使用Excel 2016、Origin 2018和R 4.0.2等软件。用Excel 2016对连续记录的温度数据进行分组和对齐时间轴。为探究垫状点地梅存活与死亡个体内部温度的差异,先对温度数据进行正态检验,确定符合正态分布后,用R 4.0.2软件中ggplot2包ggboxplot函数绘制t检验的差异箱线图。为探究垫状点地梅表面温度、内部温度、气温和土壤温度的昼夜变化规律,采用Origin 2018绘制各温度变化的折线图,为比较不同条件下垫状点地梅的保温能力是否存在显著差异,用R 4.0.2中ggpubr包和rstatix包绘制比较柱形图,用t检验的P值判断差异显著性。
2. 结果与分析
2.1 青藏高原垫状点地梅的保温机制
2.1.1 夏季垫状点地梅存活与死亡个体内部温度比较
热萨乡测得的温度数据如图2:A所示,相较于存活个体,垫状点地梅死亡个体的内部温度波动幅度更大,平均温度为19.94℃,存活个体的平均温度为19.63℃,两种个体内部温度间无显著差异。门布乡测得的温度数据如图2:B所示,存活个体和死亡个体的平均温度分别为18.18℃和17.57℃,土壤平均温度为16.45℃;存活个体内部温度高于土壤温度,且存在极显著差异(P < 0.01),但与死亡个体内部温度无显著差异(P > 0.05)。
![]() 图 2 青藏高原垫状点地梅存活与死亡个体内部温度比较A:热萨乡垫状点地梅存活与死亡个体内部温度的比较。B:门布乡垫状点地梅存活个体、死亡个体内部温度以及土壤温度的比较。 *,P < 0.05;**,P < 0.01;ns,P > 0.05。下同。Figure 2. Comparison between internal temperatures of living and dead Androsace tapete individuals on Qinghai-Tibet PlateauA: Comparison of internal temperatures of living and dead A. tapete individuals in Resa township. B: Comparison of internal temperatures of living and dead A. tapete individuals and soil temperatures in Menbu township. Same below.
图 2 青藏高原垫状点地梅存活与死亡个体内部温度比较A:热萨乡垫状点地梅存活与死亡个体内部温度的比较。B:门布乡垫状点地梅存活个体、死亡个体内部温度以及土壤温度的比较。 *,P < 0.05;**,P < 0.01;ns,P > 0.05。下同。Figure 2. Comparison between internal temperatures of living and dead Androsace tapete individuals on Qinghai-Tibet PlateauA: Comparison of internal temperatures of living and dead A. tapete individuals in Resa township. B: Comparison of internal temperatures of living and dead A. tapete individuals and soil temperatures in Menbu township. Same below.2.1.2 冬季垫状点地梅存活与死亡个体的内部温度比较
冬季垫状点地梅存活与死亡个体的内部温度、土壤温度以及附近气温的比较如图3所示。气温平均为−7.31℃,存活和死亡个体的内部平均温度分别为−4.11℃和−2.60℃,土壤平均温度为−1.06℃。分析结果表明,冬季垫状点地梅存活与死亡个体的内部温度与气温、土壤温度均有显著差异(P < 0.05),而存活与死亡个体的内部温度之间并无显著差异(P > 0.05),与夏季结果一致。
![]() 图 3 冬季气温、垫状点地梅存活个体内部温度、死亡个体内部温度以及土壤温度的比较Figure 3. Comparison among air temperature, internal temperatures of living and dead Androsace tapete individuals, and soil temperature during winter
图 3 冬季气温、垫状点地梅存活个体内部温度、死亡个体内部温度以及土壤温度的比较Figure 3. Comparison among air temperature, internal temperatures of living and dead Androsace tapete individuals, and soil temperature during winter2.2 青藏高原垫状点地梅的保温能力
2.2.1 夏季垫状点地梅表面温度和内部温度、气温、土壤温度昼夜变化
夏季3个居群的垫状点地梅温度动态变化规律如图4所示。从波动幅度来说,垫状点地梅的表面温度 > 气温 > 垫状点地梅的内部温度 > 土壤温度,保温能力则与之相反。实验中记录到的垫状点地梅表面温度最高值出现在夺底沟,为51.56℃,最低值出现在麦地卡,为−1.25℃。3个居群内,垫状点地梅的内部温度均与土壤温度相似,但从变化规律来看,随着气温的降低,其内部温度相较于土壤温度下降的更快,随着气温的上升,其内部温度相较于土壤温度上升的更快,因此其保温能力稍弱于土壤5 cm处。
![]() 图 4 夏季不同居群垫状点地梅表面和内部温度、气温、土壤温度的昼夜变化A:麦地卡居群;B:羊八井居群;C:夺底沟居群。Figure 4. Diurnal variation in surface, internal, air, and soil temperatures in different populations of Androsace tapete in summerA: Maidika population; B: Yangbajing population; C: Duodigou population.
图 4 夏季不同居群垫状点地梅表面和内部温度、气温、土壤温度的昼夜变化A:麦地卡居群;B:羊八井居群;C:夺底沟居群。Figure 4. Diurnal variation in surface, internal, air, and soil temperatures in different populations of Androsace tapete in summerA: Maidika population; B: Yangbajing population; C: Duodigou population.2.2.2 冬季垫状点地梅表面温度和内部温度、气温、土壤温度的昼夜变化
冬季垫状点地梅存活与死亡个体的表面温度、内部温度、气温和土壤温度4天的动态变化,如图5所示,垫状点地梅存活与死亡个体的温度曲线基本一致,波动幅度为:表面温度 > 气温 > 内部温度 > 土壤温度。冬季垫状点地梅存活个体表面的温度区间为−16.60℃ ~ 24.35℃,死亡个体表面的温度区间为−17.00℃ ~ 23.08℃。相较于夏季,冬季的气温显著降低,记录温度区间为−17.35℃ ~ 13.02℃,昼夜温差缩小。垫状点地梅存活个体内部温度的区间为−12.91℃ ~ 13.50℃,相较于土壤温度的变化(−6.18℃ ~ 8.47℃),其内部的温度变化更接近气温的变化,变化趋势与夏季的结果一致,其保温能力弱于土壤5 cm处。
![]() 图 5 冬季米拉山垫状点地梅表面和内部温度、气温、土壤温度的昼夜变化A:垫状点地梅存活个体;B:垫状点地梅死亡个体。Figure 5. Diurnal variation in surface, internal, air, and soil temperatures in Androsace tapete from Mila Mountain in winterA: Surviving A. tapete individuals; B: Dead A. tapete individuals.
图 5 冬季米拉山垫状点地梅表面和内部温度、气温、土壤温度的昼夜变化A:垫状点地梅存活个体;B:垫状点地梅死亡个体。Figure 5. Diurnal variation in surface, internal, air, and soil temperatures in Androsace tapete from Mila Mountain in winterA: Surviving A. tapete individuals; B: Dead A. tapete individuals.2.2.3 低温处理对垫状点地梅表面及内部温度的影响
低温处理后,各温度的动态变化如图6所示。撒下碎冰时,垫状点地梅的表面温度、内部温度以及土壤温度迅速下降,降温速率为表面 > 内部 >土壤。当碎冰完全融化时开始升温,升温速率为表面 > 内部 > 土壤,与昼夜温度变化的结果一致,即垫状点地梅的保温能力稍弱于土壤5 cm处。整个过程中,垫状点地梅的内部温度均低于土壤温度,但最低温度仍维持在12℃以上。
![]() 图 6 模拟冰雹处理对垫状点地梅表面及内部温度的影响Figure 6. Effect of three simulated hail treatments on surface and internal temperatures in Androsace tapete
图 6 模拟冰雹处理对垫状点地梅表面及内部温度的影响Figure 6. Effect of three simulated hail treatments on surface and internal temperatures in Androsace tapete2.2.4 不同条件下垫状点地梅保温能力的比较
为探究垫状点地梅内部温度与气温间差异的显著性,对日间和夜间各居群垫状点地梅的内部温度与气温做差异性分析(夏季日出到日落时间为7:00 – 21:00,冬季日出到日落时间为8:30 – 19:00),结果如图7所示。在日间不同海拔的居群中,除麦地卡和羊八井外,其他居群的垫状点地梅内部温度与气温均存在极显著差异(P < 0.01),且在夏季其内部平均温度略低于气温,在冬季则高于气温。在夜间,各居群的垫状点地梅内部温度与气温均存在极显著差异(P < 0.01),且其内部温度远高于气温。无论是在日间还是夜间,总体上垫状点地梅内部温度的变化要显著小于气温的变化。
![]() 图 7 垫状点地梅内部温度与气温的比较A:日间垫状点地梅内部温度与气温的比较。B:夜间垫状点地梅内部温度与气温的比较。DDG:夏季夺底沟居群;MDK:夏季麦地卡居群;MLS-A:冬季米拉山存活个体样品;MLS-D:冬季米拉山死亡个体样品; YBJ:夏季羊八井居群。Figure 7. Comparison between internal temperature of Androsace tapete and air temperatureA: Comparison between internal temperature of A. tapete and air temperature during daytime. B: Comparison between internal temperature of A. tapete and air temperature at night. “DDG”: Summer Duodigou population; “MDK”: Summer Maidika population; “MLS-A”: Samples of alive individuals from Mila Mountain in winter; “MLS-D”: Samples of dead individuals from Mila Mountain in winter; “YBJ”: Summer Yangbajing population.
图 7 垫状点地梅内部温度与气温的比较A:日间垫状点地梅内部温度与气温的比较。B:夜间垫状点地梅内部温度与气温的比较。DDG:夏季夺底沟居群;MDK:夏季麦地卡居群;MLS-A:冬季米拉山存活个体样品;MLS-D:冬季米拉山死亡个体样品; YBJ:夏季羊八井居群。Figure 7. Comparison between internal temperature of Androsace tapete and air temperatureA: Comparison between internal temperature of A. tapete and air temperature during daytime. B: Comparison between internal temperature of A. tapete and air temperature at night. “DDG”: Summer Duodigou population; “MDK”: Summer Maidika population; “MLS-A”: Samples of alive individuals from Mila Mountain in winter; “MLS-D”: Samples of dead individuals from Mila Mountain in winter; “YBJ”: Summer Yangbajing population.不同条件下垫状点地梅保温能力的比较如图8:A所示。不同海拔居群中,夺底沟与羊八井的垫状点地梅居群保温能力存在显著差异(P < 0.05),其余不同生存状态和不同季节的垫状点地梅居群之间的保温能力均无显著差异。不同条件下垫状点地梅与5 cm土层的保温能力比较如图8:B所示。5 cm土层的保温能力均值均高于垫状点地梅,在夺底沟和米拉山两个样点中,5 cm土层的保温能力显著高于垫状点地梅(P < 0.05)。综上,海拔差、存活与否以及季节的更替不会显著影响垫状点地梅的保温能力,且垫状点地梅的保温能力稍弱于5 cm的土层。
![]() 图 8 不同条件下垫状点地梅保温能力的比较A:不同条件下垫状点地梅的保温能力。B:不同条件下垫状点地梅与5 cm 土层的保温能力。DDG:夺底沟居群;MDK:麦地卡居群;MLS-A:米拉山存活个体样品;MLS-D:米拉山死亡个体样品;YBJ:羊八井居群。Figure 8. Comparison of heat-retaining capacity of Androsace tapete under different conditionsA: Heat-retaining capacity of A. tapete under different conditions. B: Heat-retaining capacity of A. tapete and 5-cm soil layer under different conditions. “DDG”: Duodigou population; ‘MDK”: Maidika population; “MLS-A”: Samples of alive individuals from Mila Mountain; “MLS-D”: Samples of dead individuals from Mila Mountain; “YBJ”: Yangbajing population.
图 8 不同条件下垫状点地梅保温能力的比较A:不同条件下垫状点地梅的保温能力。B:不同条件下垫状点地梅与5 cm 土层的保温能力。DDG:夺底沟居群;MDK:麦地卡居群;MLS-A:米拉山存活个体样品;MLS-D:米拉山死亡个体样品;YBJ:羊八井居群。Figure 8. Comparison of heat-retaining capacity of Androsace tapete under different conditionsA: Heat-retaining capacity of A. tapete under different conditions. B: Heat-retaining capacity of A. tapete and 5-cm soil layer under different conditions. “DDG”: Duodigou population; ‘MDK”: Maidika population; “MLS-A”: Samples of alive individuals from Mila Mountain; “MLS-D”: Samples of dead individuals from Mila Mountain; “YBJ”: Yangbajing population.3. 讨论
3.1 垫状点地梅的保温作用
本研究分析了不同条件下垫状点地梅的保温能力,发现总体上不同海拔、不同季节和不同生存状下的垫状点地梅,其保温能力并无显著差异,其内部温度的变化要显著小于气温的变化。日间,金属探头所记录的温度会略高于周围气温,其差异程度取决于探头的材质、大小、形状和太阳直射面积等因素。本研究所用的探头长5 cm,直径2 mm,受太阳辐射直接增温的效应较小[30, 31]。更准确的夜间温度数据表明,垫状点地梅内部温度的变幅远小于气温的变幅。且在高海拔的两个居群和冬季米拉山居群中,其内部温度均显著高于气温,与日间温度变化趋势相同,即垫状点地梅能有效缓冲气温变化对垫状体的影响,以此减轻夏季日间高温和冬季或夜间低温的胁迫,这也与夏季和冬季的昼夜温度变化曲线一致。在夏季日间,垫状点地梅的内部温度显著高于土壤温度,其表面温度也远高于附近气温,能够在生长季节完成有效积温。垫状点地梅的死亡个体虽然失去了生理活性,但仍然维持着相同的结构特征,因此推测垫状点地梅的保温作用可能与自身代谢活动、冷驯化、海拔差异等因素无关,而主要是与其球状或椭球状、外部紧密内部松散、比表面积小等的结构特性有关,与之前的研究结果相吻合[32]。
垫状点地梅内部温度与土壤温度的变化相似,温度波动幅度略大于土壤温度,保温能力略弱于5 cm的土层。垫状点地梅与空气接触的面积较大,且突出于地表,其附近土壤的质地紧密、透气性较差,因此垫状点地梅比土壤更易受气温变化的影响,这与在垫状植物Diapensia lapponica L.中的研究结果相一致[13]。与垫状植物Silence acaulis L.表面温度增加或下降的速率远大于气温增加或下降速率相似[12, 33],垫状点地梅的表面温度波动幅度和频率更大,易受太阳辐射、云、风和雨等外界环境影响,在晴天可以迅速升温,积蓄热能,本研究中记录到的最高温度甚至超过了50℃。在夜间温度骤降时,垫状点地梅的表面温度会降至露点以下,有利于空气中的水分在其表面凝结,从而有利于其在干旱环境中吸收水分。垫状植物表面的昼夜温度变化幅度远高于内部[13],相较于垫状植物Azorella compacta Phil.的42℃[34],垫状点地梅表面昼夜日温差最高达44.63℃,而内部的最大温差仅19.69℃。
本研究夏季数据表明,夜间较长时间的低温也可将垫状点地梅“冻透”,其内部温度降至与气温一致,甚至更低。冬季数据则表明,在气温下降至−17℃时,垫状点地梅内部的温度能维持在−12.91℃以上,较于夏季内部温度与气温的差值有所增加,但内部温度的变化趋势仍然相似,弱于5 cm处土层的保温作用。因此,垫状点地梅的保温能力并没有预期的强,但其垫状结构缓冲了环境温度变化带来的高温或低温胁迫的影响,是其能够在昼夜温差、季节温度变化巨大的高海拔地区生存的重要因素。此外,今后应关注垫状点地梅形成其垫状结构的适应性进化机制,以及其叶片抗高温、耐低温的内部分子机制,如全球变暖使青藏高原高海拔植物的蒸腾失水的压力增加,相较于温度上升对垫状植物的影响,这可能是触发高山植物将叶片和茎进化为更小、更密集的垫状适应性结构的关键因素[22]。
3.2 全球变暖及冷驯化对垫状点地梅的影响
垫状点地梅表面的叶片可以在一昼夜中承受近50℃的温度变化,理论上来说,全球变暖导致的温度变化幅度不足以对其生存造成严重威胁。但它生长在海拔4300 ~ 5300 m的平缓山口或砾质山坡的生境中[35],而高山山顶对气候变化特别敏感,高海拔的山顶区域正经历着最高的变暖率[36, 37]。事实上,即便温度的微小变化也可能导致垫状点地梅生境的温度上限升高、下限降低,从而超过其最大耐受程度,同时还会引起蒸腾失水的压力增加[38]。在过去45年中,青藏高原极端高温和低温的出现频率增加已被证实[39],像垫状点地梅这种适应了高海拔极端气候条件的植物,即便是有着较高迁移能力的种子植物,但在通过迁移来适应气候变化的选择仍然很少[40, 41],尤其是生长在山顶处的居群。不仅如此,全球变暖可能还会导致与垫状点地梅竞争的植物发生改变,如高山嵩草Carex parvula O. Yano等会进一步挤压其生态位[42]。
众所周知,植物的冷驯化是一个渐变而长期的过程,如秋季过渡到冬季。经冷驯化后的植物将获得更高的抗寒能力,能有效地抵御低温冻伤。在青藏高原高海拔的极端环境中,不仅昼夜温差大,而且即便是夏季的晴天冰雹等恶劣天气也经常发生,这对植物适应温度骤降是一个严峻的挑战。本研究模拟突发的冰雹天气,对垫状点地梅进行撒冰的整个过程中,其内部的最低温度仍能维持在12℃以上。随着碎冰的融化,垫状点地梅表面缝隙中的枯枝败叶等凋落物吸水膨胀,使其表面结构较未撒冰前变得更加紧密。因此,垫状点地梅能够在无冷驯化的条件下,利用自身的保温能力和致密强韧的结构,隔绝冰雹带来的低温冻伤和机械损伤,以适应高海拔多变的天气。另外,在冬季经自然冷驯化后,垫状点地梅的保温能力并未显著增强,垫状结构只能使其核心温度高于气温4℃左右,内部温度最低能达到−12.91℃,推测此时其对严寒的抵御可能与提高细胞膜脂中的类脂和脂肪酸成分的不饱和度[43]、某些酶类通过变构调节克服低温钝化[44]、提高抗氧化酶活性[45]、增加渗透调节物质浓度[46]等其他内部分子机制有关。
4. 结论
垫状点地梅外部致密内部疏松的垫状结构使其具有一定的保温作用,其保温能力略弱于5 cm的土层,突出于地面的球形或椭球形表面能在日照强烈时有效地积累热量,较为松散的内部在外界温度骤降时可缓慢释放积蓄的热量,显著缓冲了高原昼夜剧烈的温差。内部较高的温度保护了嫩叶、枝芽和花苞,为植株夏季进行营养生长和生殖生长提供保障。夏季,面对突发的冰雹天气,垫状点地梅较高的表面温度能使散落在其表面的冰雹加速融化,叶片和茎间缝隙中的凋落物吸水膨胀,变得更加紧密,能有效抵御冰雹带来的冻伤和机械损伤。冬季,经冷驯化后的垫状点地梅,其保温能力并未显著增强,但其内部温度仍显著高于外界环境温度,此时植株的叶和茎枯萎,以休眠状态过冬。
-
![]()
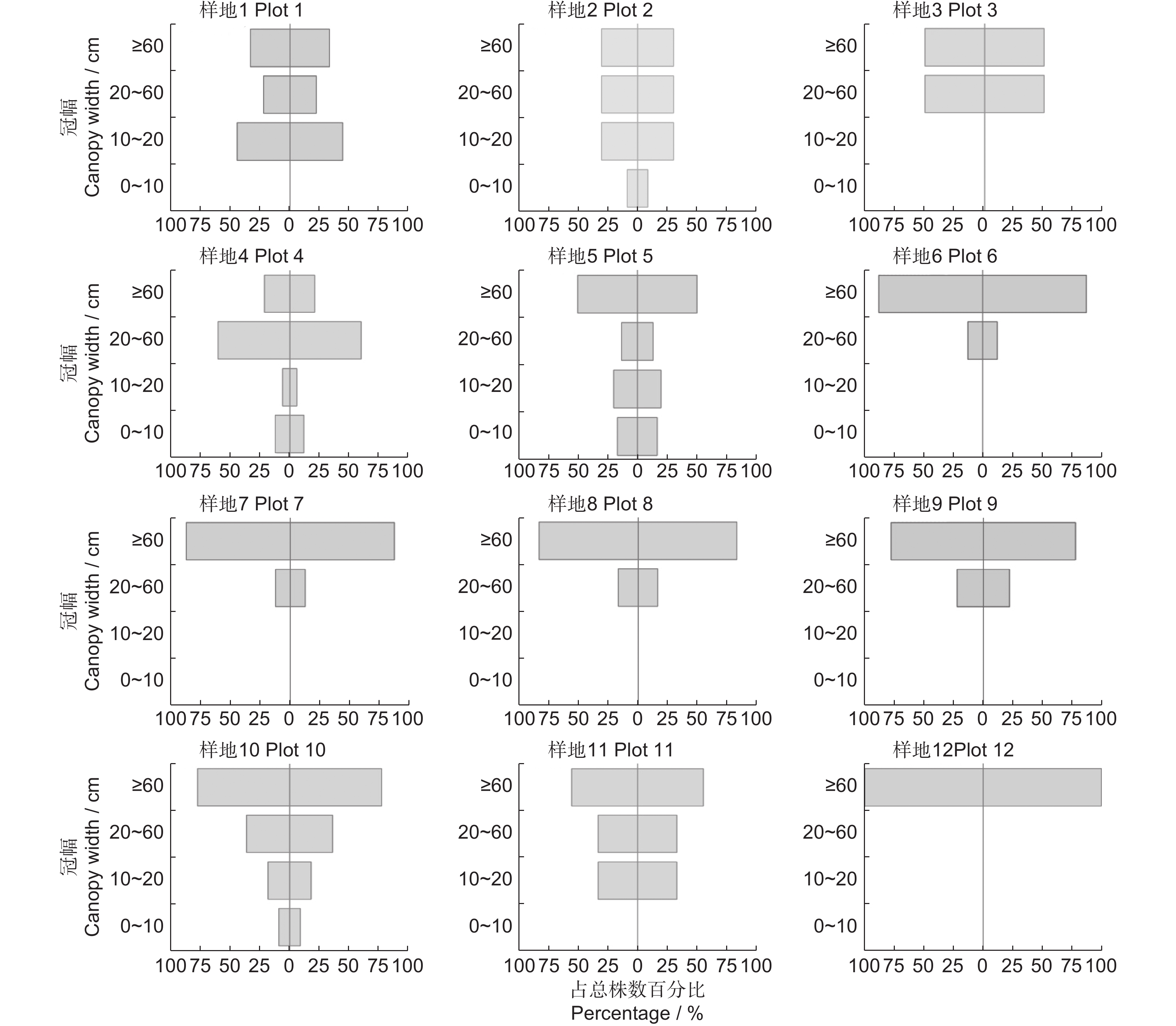
图 1 不同样方的新疆沙冬青种群年龄结构
Figure 1. Age structure of Ammopiptanthus nanus populations in different plots
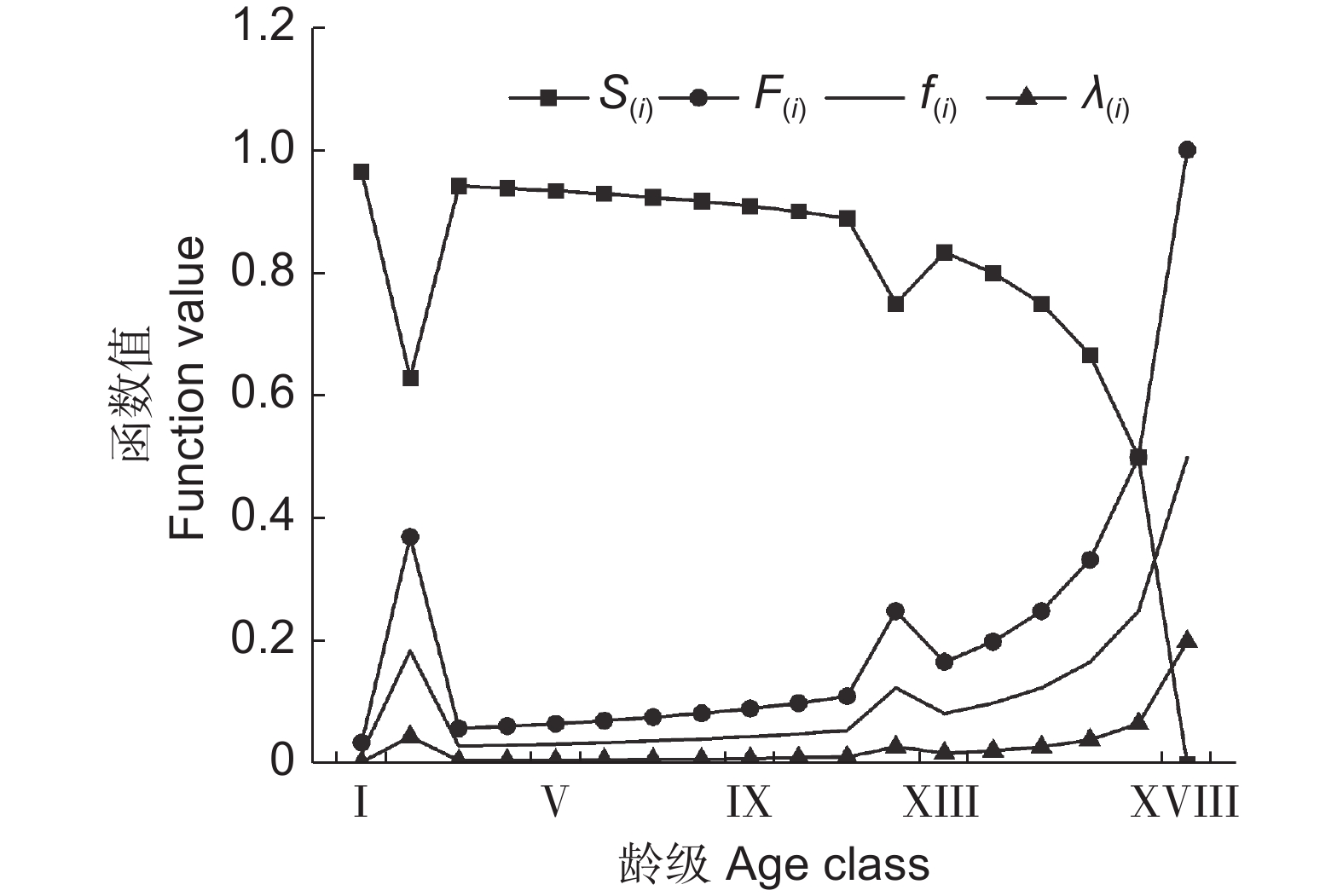
![]()
图 4 新疆沙冬青种群生存函数曲线
S(i):生存率函数;F(i):累积死亡率函数;f(ti):死亡密度;λ(ti):危险率函数。
Figure 4. Survival function curves of Ammopiptanthus nanus populations
S(i): Survival rate function; F(i): Cumulative mortality rate function; f(ti): Mortality density rate function; λ(ti): Hazard rate function.
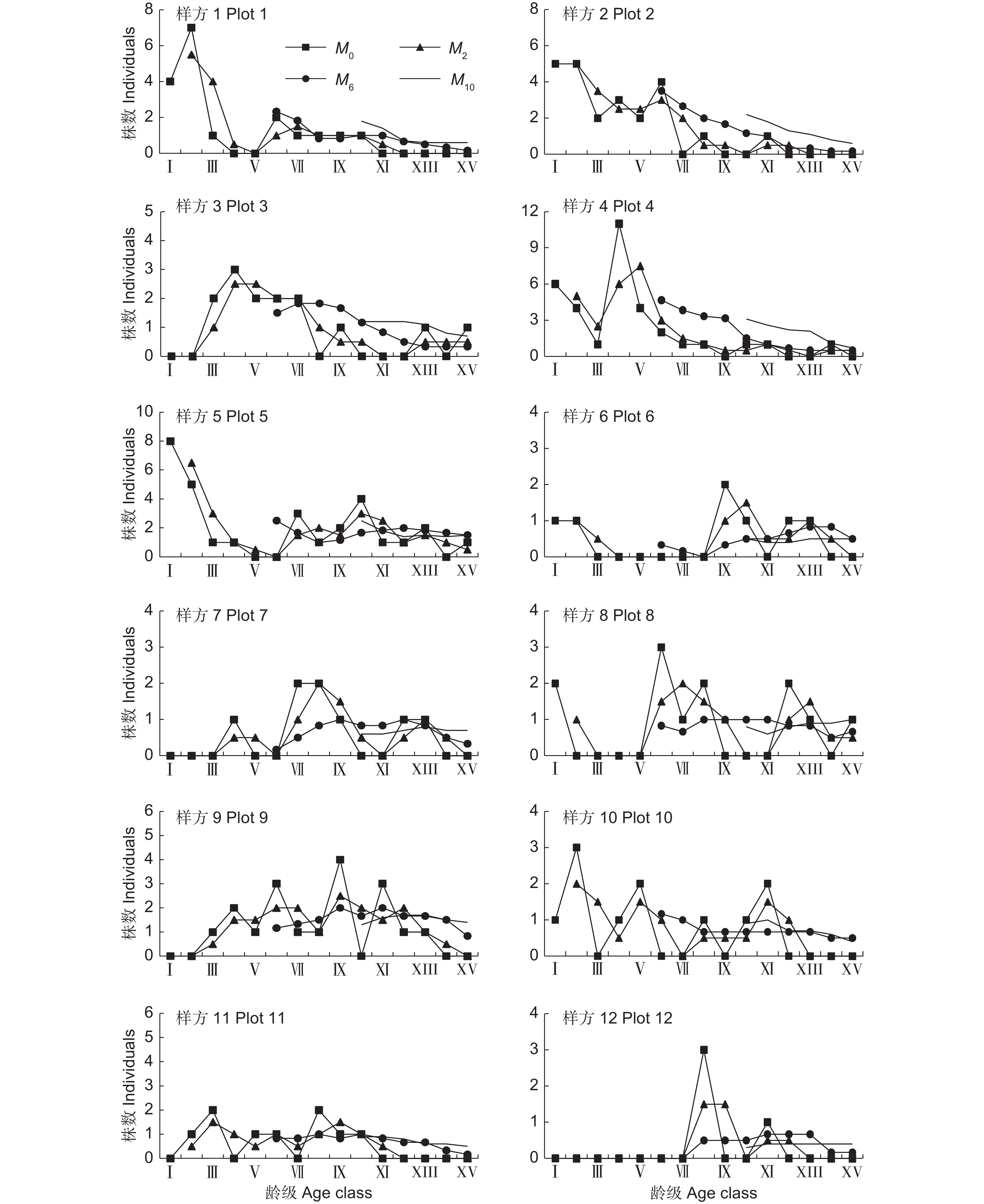
![]()
图 5 新疆沙冬青种群动态的时间序列分析
M0:各龄级现有株数;M2:各龄级在未来2个龄级时间后的株数;M6:各龄级在未来6个龄级时间后的株数;M10:各龄级在未来10个龄级时间后的株数。
Figure 5. Time series analysis of population dynamics of Ammopiptanthus nanus
M0: Number of individuals in each age group; M2: Number of individuals in each age class after the next two age classes; M6: Number of individuals in each age class after the next six age classes; M10: Number of individuals in each age class after the next 10 age classes.
表 1 12个样方的地理分布
Table 1 Geographical information of 12 community plots of Ammopiptanthus nanus
样方号
No. of plot纬度
Latitude经度
Longitude海拔
Altitude / m坡向
Aspect1 39°48′N 75°18′E 2 395 东 2 39°48′N 75°18′E 2 395 东 3 39°48′N 75°18′E 2 405 西北 4 39°49′N 75°18′E 2 434 西 5 39°39′N 75°01′E 2 066 西 6 39°39′N 75°01′E 2 060 西 7 39°28′N 74°54′E 2 395 西 8 39°28′N 74°54′E 2 395 东 9 39°28′N 74°54′E 2 393 东北 10 39°28′N 74°54′E 2 393 西南 11 39°49′N 75°35′E 2 032 东北 12 39°49′N 75°35′E 2 032 东南  下载: 导出CSV
下载: 导出CSV
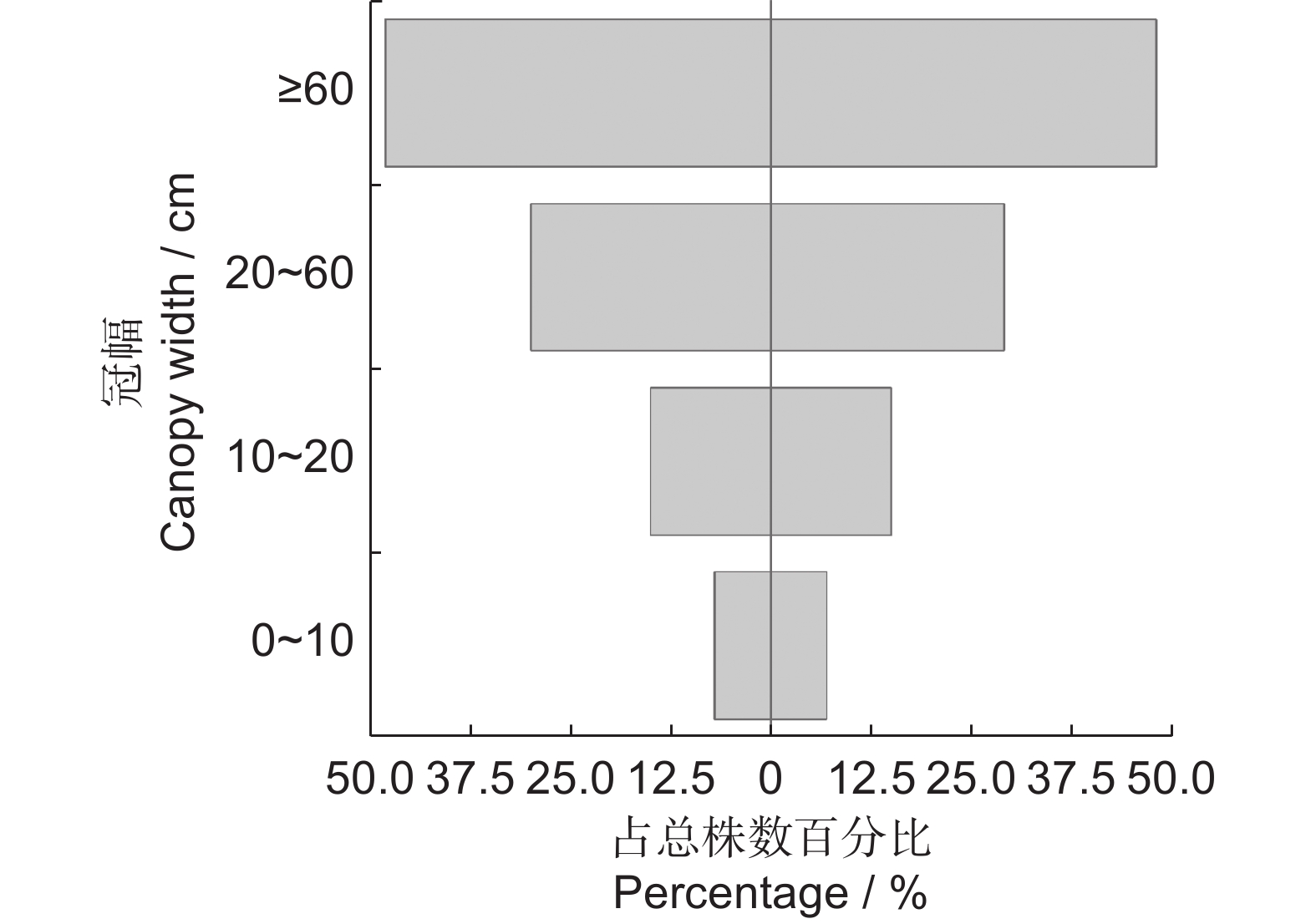
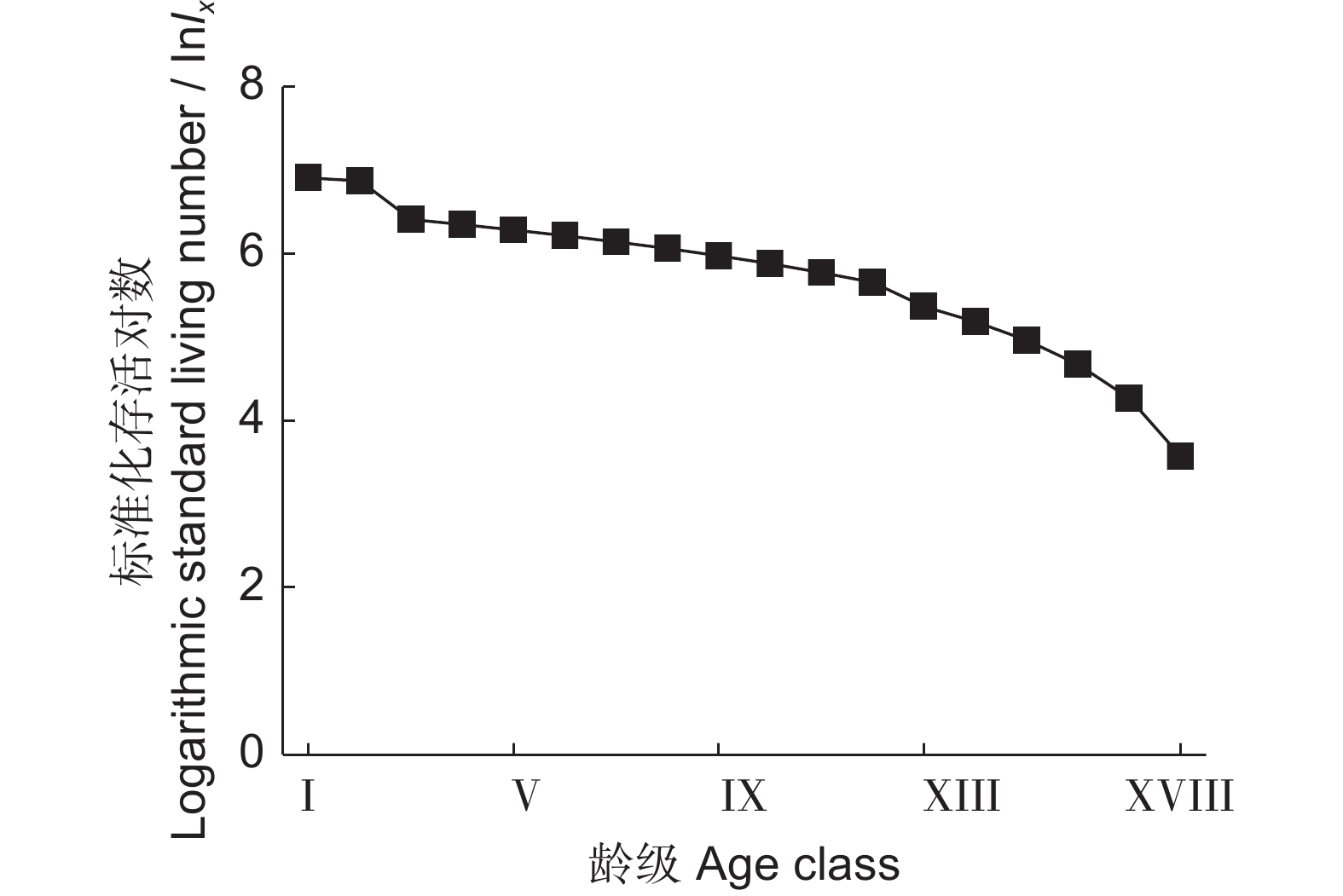
表 2 新疆沙冬青静态生命表
Table 2 Static life table of Ammopiptanthus nanus populations
龄级
Age class冠幅
Canopy width / cmax ax' lx lnlx dx Lx Tx qx ex Kx Ⅰ 0~10 13 28 1 000 6.91 36 982 6 679 0.04 6.68 0.04 Ⅱ 10~20 28 27 964 6.87 357 786 5 697 0.37 5.91 0.46 Ⅲ 20~30 17 17 607 6.41 36 589 4 911 0.06 8.09 0.06 Ⅳ 30~40 7 16 571 6.35 36 554 4 322 0.06 7.56 0.06 Ⅴ 40~50 17 15 536 6.28 36 518 3 768 0.07 7.03 0.07 Ⅵ 50~60 15 14 500 6.21 36 482 3 250 0.07 6.50 0.07 Ⅶ 60~70 12 13 464 6.14 36 446 2 768 0.08 5.96 0.08 Ⅷ 70~80 12 12 429 6.06 36 411 2 322 0.08 5.42 0.09 Ⅸ 80~90 10 11 393 5.97 36 375 1 911 0.09 4.86 0.10 Ⅹ 90~100 13 10 357 5.88 36 339 1 536 0.10 4.30 0.11 Ⅺ 100~110 10 9 321 5.77 36 304 1 197 0.11 3.72 0.12 Ⅻ 110~120 10 8 286 5.65 71 250 893 0.25 3.13 0.29 ⅩⅢ 120~130 5 6 214 5.37 36 196 643 0.17 3.00 0.18 ⅩⅣ 130~140 6 5 179 5.18 36 161 447 0.20 2.50 0.22 ⅩⅤ 140~150 4 4 143 4.96 36 125 286 0.25 2.00 0.29 ⅩⅥ 150~160 4 3 107 4.67 36 89 161 0.33 1.50 0.41 ⅩⅦ 160~170 3 2 71 4.27 36 54 72 0.50 1.00 0.69 ⅩⅧ ≥170 1 1 36 3.58 − 18 18 − 0.50 − 注:ax为相应龄级的个体数;ax'为匀滑处理后的个体数;lx为x龄级标准化存活个体数,lx=ax/a0×1 000;dx为从x到x+1龄级间标准化的死亡数,dx=lx–lx+1;qx为从x到x+1龄级间的个体死亡率,qx=dx/lx;Lx为从x到x+1龄级的平均存活个体数,Lx=(lx/lx+1)/2;Tx为大于等于x龄级的各龄级存活数,Tx=∫∞XLx;ex为x龄级平均期望寿命,ex=Tx/lx;Kx为致死率,Kx=ln(lx)−ln(lx+1)。 Notes: ax represents the number of individuals of corresponding age class; ax' represents the number of individuals after smoothing; lx is the number of standardized surviving individuals of x age class, lx=ax/a0×1 000; dx represents the standardized death number from x age class to x+1, dx=lx-lx+1; qx represents the mortality rates from x age class to x+1, qx=dx/lx; Lx represents the average number of surviving individuals from x age class to x+1, Lx=(lx/lx+1)/2; Tx is the number of survivors of all ages greater than or equal to the x age class, Tx=∫∞XLx; ex represents the average life expectancy of the x age class, ex=Tx/lx; Kx represents the death rate, Kx=ln(lx)−ln(lx+1).
下载: 导出CSV
-
[1] Li W,Zhang GF. Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae)[J]. Flora,2015,212:10−18. doi: 10.1016/j.flora.2015.02.002
[2] Zhang QY,Zhang XX,Liang ZX,Farooq A,Luo JR,et al. Population dynamics of the endangered plant Paeonia qiui (Paeoniaceae)[J]. Pak J Bot,2020,52(4):1353−1359.
[3] 牛翠娟,娄安如,孙儒泳,李庆芬. 基础生态学[M]. 3版. 北京:高等教育出版社,2015:150−151. [4] Chhetri PK,Bista R,Cairns DM. Population structure and dynamics of Abies spectabilis at treeline ecotone of Barun Valley,Makalu Barun National Park,Nepal[J]. Acta Ecol Sin,2016,36(4):269−274. doi: 10.1016/j.chnaes.2016.05.003
[5] 刘建泉,罗永寿,吕海元. 不同封育季节放牧干扰对青海云杉种群结构和动态的影响[J]. 草业科学,2012,29(6):983−988. Liu JQ,Luo YS,Lü HY. Structure and dynamics of Picea crassifolia populations with graze disturbance in different seasons of enclosed forest[J]. Pratacultural Science,2012,29(6):983−988.
[6] Yuan GY,Guo QQ,Xie N,Zhang YQ,Luo SQ,et al. Population dynamics and protection strategies of Litsea coreana Levl. var. sinensis with Guizhou Province in China as an example[J]. J Mt Sci,2023,20(4):1090−1101. doi: 10.1007/s11629-022-7429-z
[7] Feng L,Li N,Yang W,Li Y,Wang CM,et al. Analyses of mitochondrial genomes of the genus Ammopiptanthus provide new insights into the evolution of legume plants[J]. Plant Syst Evol,2019,305(5):385−399. doi: 10.1007/s00606-019-01578-2
[8] 段义忠,张凯. 沙冬青属植物叶绿体基因组对比和系统发育分析[J]. 西北植物学报,2020,40(8):1323−1332. doi: 10.7606/j.issn.1000-4025.2020.08.1323 Duan YZ,Zhang K. Comparative analysis and phylogenetic evolution of the complete chloroplast genome of Ammopiptanthus[J]. Acta Botanica Boreali-Occidentalia Sinica,2020,40(8):1323−1332. doi: 10.7606/j.issn.1000-4025.2020.08.1323
[9] 中国科学院中国植物志编辑委员会. 中国植物志:第42卷,第2分册[M]. 北京:科学出版社,1998:395−397. [10] 郝文芳,周禧琳,王海珍,刘艳萍. 濒危植物小沙冬青研究进展[J]. 植物科学学报,2019,37(1):109−115. doi: 10.11913/PSJ.2095-0837.2019.10109 Hao WF,Zhou XL,Wang HZ,Liu YP. Research progress on rare and endangered plant Ammopiptanthus nanus[J]. Plant Science Journal,2019,37(1):109−115. doi: 10.11913/PSJ.2095-0837.2019.10109
[11] Ji TF,Li J,Liang CH. The chemical constituents of the twigs of Ammopiptanthus nanus[J]. J Asian Nat Prod Res,2013,15(4):332−336. doi: 10.1080/10286020.2012.762765
[12] Wei HH,Wu P,Ge XJ,Liu MF,Wei XY. Chemical constituents of the seeds of Ammopiptanthus (Leguminosae) and their systematic and ecological significance[J]. Biochem Syst Ecol,2007,35(5):274−280. doi: 10.1016/j.bse.2006.10.015
[13] 傅立国. 中国植物红皮书-稀有濒危植物:第一册[M]. 北京:科学出版社,1991:370−371. [14] 尹林克. 新疆珍稀濒危特有高等植物[M]. 乌鲁木齐:新疆科学技术出版社,2006:66. [15] 杨期和,葛学军,叶万辉,邓雄,廖富林. 矮沙冬青种子特性和萌发影响因素的研究[J]. 植物生态学报,2004,28(5):651−656. doi: 10.3321/j.issn:1005-264X.2004.05.010 Yang QH,Ge XJ,Ye WH,Deng X,Liao FL. Characteristics of Ammopiptanthus nanus seed and factors affecting its germination[J]. Acta Phytoecologica Sinica,2004,28(5):651−656. doi: 10.3321/j.issn:1005-264X.2004.05.010
[16] 高志海,刘生龙,仲述军,王理德. 矮沙冬青引种栽培试验研究[J]. 甘肃林业科技,1995,20(1):28−31,34. Gao ZH,Liu SL,Zhong SJ,Wang LD. Experiments on introduction and culture of Ammopiptanthus nanus[J]. Journal of Gansu Forestry Science and Technology,1995,20(1):28−31,34.
[17] 李新蓉,谭敦炎. 新疆沙冬青(Ammopiptanthus nanus)的开花物候与环境的关系[J]. 中国沙漠,2007,27(4):572−578. doi: 10.3321/j.issn:1000-694X.2007.04.008 Li XR,Tan DY. Relationships between flowering phenology of Ammopiptanthus nanus (Fabaceae) and its environment[J]. Journal of Desert Research,2007,27(4):572−578. doi: 10.3321/j.issn:1000-694X.2007.04.008
[18] Cheng YS,Bai LP,Zhang L,Chen G,Fan JG,et al. Identification and characterization of AnICE1 and AnCBFs involved in cold tolerance from Ammopiptanthus nanus[J]. Plant Physiol Biochem,2021,168:70−82. doi: 10.1016/j.plaphy.2021.09.030
[19] Li AR,Ma M,Li HT,He SF,Wang SG. Genetic diversity and population differentiation of a Chinese endangered plant Ammopiptanthus nanus (M. Pop.) Cheng f.[J]. Genes,2023,14(5):1020. doi: 10.3390/genes14051020
[20] 杨鹏鹏. 乌恰县近60年气象要素变化分析[J]. 西北水电,2018(6):13−16. doi: 10.3969/j.issn.1006-2610.2018.06.003 Yang PP. Analysis on meteorological factor variation in recent 60 years in Wuqia County[J]. Northwest Hydropower,2018(6):13−16. doi: 10.3969/j.issn.1006-2610.2018.06.003
[21] 靳虎甲,王继和,马全林. 乌兰布和沙漠沙冬青种群结构和空间分布[J]. 西北植物学报,2010,30(11):2300−2306. Jin HJ,Wang JH,Ma QL. Size structure and distribution pattern of Ammopiptanthus mongolicus population in Ulanbuh desert[J]. Acta Botanica Boreali-Occidentalia Sinica,2010,30(11):2300−2306.
[22] 刘虎俊,尉秋实,王继和,廖空太,马瑞,等. 甘肃北砸山沙冬青种群的空间分布格局[J]. 干旱区研究,2009,26(6):865−871. Liu HJ,Wei QS,Wang JH,Liao KT,Ma R,et al. Spatial distribution patterns of the population of Ammopiptanthus mongolicus in the Beiza Mountain,China[J]. Arid Zone Research,2009,26(6):865−871.
[23] 尉秋实,王继和,李昌龙,庄光辉,陈善科. 不同生境条件下沙冬青种群分布格局与特征的初步研究[J]. 植物生态学报,2005,29(4):591−598. doi: 10.3321/j.issn:1005-264X.2005.04.010 Wei QS,Wang JH,Li CL,Zhuang GH,Chen SK. A preliminary study on the distribution patterns and characeristics of Ammopiptanthus mongolicus populations in different desert environments[J]. Acta Phytoecologica Sinica,2005,29(4):591−598. doi: 10.3321/j.issn:1005-264X.2005.04.010
[24] 杨彪,张全建,龚旭,段晨松,张远彬. 雅砻江冬麻豆 (Salweenia bouffordiana) 种群结构与动态特征[J]. 生态学报,2020,40(4):1184−1194. Yang B,Zhang QJ,Gong X,Duan CS,Zhang YB. Population structure and dynamic characteristics of Salweenia bouffordiana[J]. Acta Ecologica Sinica,2020,40(4):1184−1194.
[25] 赵鹏,徐先英,张裕年,段晓峰,邱晓娜,等. 民勤绿洲荒漠过渡带人工梭梭种群年龄结构及动态[J]. 生态学报,2023,43(14):6069−6079. Zhao P,Xu XY,Zhang YN,Duan XF,Qiu XN,et al. Age structure and its dynamics of artificial Haloxylon ammodendron population in Minqin oasis-desert ecotone[J]. Acta Ecologica Sinica,2023,43(14):6069−6079.
[26] 江洪. 云杉种群生态学[M]. 北京:中国林业出版社,1992:8−30. [27] Hett JM,Loucks OL. Age structure models of balsam fir and eastern hemlock[J]. J Ecol,1976,64(3):1029−1044. doi: 10.2307/2258822
[28] 王世彤,李玲丽,徐耀粘,谷志容,王业清,等. 极小种群野生植物长果安息香种群结构与动态特征[J]. 植物科学学报,2022,40(5):627−636. doi: 10.11913/PSJ.2095-0837.2022.50627 Wang ST,Li LL,Xu YZ,Gu ZR,Wang YQ,et al. Population structure and dynamic characteristics of extremely small populations of Changiostyrax dolichocarpa C. J. Qi[J]. Plant Science Journal,2022,40(5):627−636. doi: 10.11913/PSJ.2095-0837.2022.50627
[29] 张文辉,祖元刚,刘国彬. 十种濒危植物的种群生态学特征及致危因素分析[J]. 生态学报,2002,22(9):1512−1520. doi: 10.3321/j.issn:1000-0933.2002.09.021 Zhang WH,Zu YG,Liu GB. Population ecological characteristics and analysis on endangered cause of ten endangered plant species[J]. Acta Ecologica Sinica,2002,22(9):1512−1520. doi: 10.3321/j.issn:1000-0933.2002.09.021
[30] 何恒斌,郝玉光,丁琼,贾桂霞. 沙冬青植物群落特征及其根瘤多样性研究[J]. 北京林业大学学报,2006,28(4):123−128. doi: 10.3321/j.issn:1000-1522.2006.04.023 He HB,Hao YG,Ding Q,Jia GX. Characteristics of plant community of Ammopiptanthus mongolicus and the diversity of its nodules[J]. Journal of Beijing Forestry University,2006,28(4):123−128. doi: 10.3321/j.issn:1000-1522.2006.04.023
[31] 潘伯荣,余其立,严成. 新疆沙冬青生态环境及渐危原因的研究[J]. 植物生态学与地植物学学报,1992,16(3):276−282. Pan BR,Yu QL,Yan C. Study for the ecological environment and vulnerable reasons of the Ammopiptanthus nanus[J]. Acta Phytoecologica et Geobotanica Sinica,1992,16(3):276−282.
[32] 潘伯荣,谢静霞,段士民. 新疆沙冬青果实虫蚀对其自然繁衍的影响[C]//第二届中国甘草学术研讨会暨第二届新疆植物资源开发、利用与保护学术研讨会论文摘要集. 石河子:中国植物学会,2004:112. [33] 黄冬柳,梁士楚,姜勇. 桂林岩溶石山青冈种群数量动态[J]. 生态学杂志,2019,38(3):663−671. Huang DL,Liang SC,Jiang Y. Quantitative dynamics of Cyclobalanopsis glauca population in Guilin karst mountainous areas[J]. Chinese Journal of Ecology,2019,38(3):663−671.
[34] 吉也,曹孟岩,白楚锋,金林琦,李畅,李萍. 峨眉山桫椤种群结构与动态特征[J]. 西北植物学报,2019,39(3):543−551. doi: 10.7606/j.issn.1000-4025.2019.03.0543 Ji Y,Cao MY,Bai CF,Jin LQ,Li C,Li P. Population structure and dynamics of Alsophila spinulosa in Mount Emei[J]. Acta Botanica Boreali-Occidentalia Sinica,2019,39(3):543−551. doi: 10.7606/j.issn.1000-4025.2019.03.0543









计量
- 文章访问数: 154
- HTML全文浏览量: 43
- PDF下载量: 15
