
Comparative study on the community characteristics and population dynamics of Bruguiera sexangula (Lour.) Poiret at different intertidal elevations in Dongzhaigang National Nature Reserve
-
摘要:
为了解不同生境下海莲(Bruguiera sexangula (Lour.) Poiret)的群落特征和种群动态变化,合理利用与保护现有红树资源,对海南东寨港国家级自然保护区不同潮位下海莲的群落特征、年龄结构、分布格局、静态生命表以及生存函数等进行了研究。结果显示:(1)海莲为群落优势树种,群落的物种多样性指数、均匀度指数以及林分密度随潮位升高逐渐增加,而海莲的重要值、胸径、树高以及冠幅随潮位升高逐渐降低;(2)海莲种群在空间上呈现聚集分布格局,不同潮位的龄级结构分布不同,低潮位种群为“凸”型而中高潮位种群为倒“J”型;(3)海莲种群的存活曲线在不同潮位上存在差异,低潮位植株死亡率在各龄级中相近,为Deevey-Ⅱ型,中高潮位植株死亡率在低龄级阶段高,在高龄级阶段相对稳定,为Deevey-Ⅲ型;(4)不同潮位海莲种群的生存率和累计死亡率函数平衡点均在幼苗阶段,第Ⅴ龄级后各潮位生存函数曲线的变化幅度变小。因此,不同潮位下海莲的群落特征和种群动态有明显差异。研究结果为该区域红树植物的保护及优化管理提供了重要的基础数据。
Abstract:Understanding how intertidal elevation influences the community characteristics and population dynamics of Bruguiera sexangula (Lour.) Poiret is essential for the effective conservation and sustainable management of mangrove ecosystems. This study investigated variations in community characteristics, age structure, spatial distribution pattern, static life-table parameters, and survival functions of B. sexangula along an intertidal gradient in the Hainan Dongzhaigang National Nature Reserve. Results showed the following key findings: (1) B. sexangula was the dominant species in the community. Species diversity, Pielou index, and stand density increased with increasing intertidal elevation, while the importance value, diameter at breast height, tree height, and crown width of B. sexangula decreased. (2) The spatial distribution pattern of B. sexangula exhibited significant clumping. The diameter distribution exhibited a convex-shaped pattern at low intertidal elevations and an inverted J-shaped pattern at both mid and high intertidal elevations. (3) Survival curves varied with intertidal elevation. Low intertidal zones exhibited a Deevey Type Ⅱ curve, while mid and high intertidal elevations showed Deevey Type Ⅲ curves. (4) The equilibrium points of survival and cumulative mortality functions occurred during the seedling stage across all intertidal elevations, and the magnitude of changes in the survival function curves at each intertidal elevation decreased after age Ⅴ. These findings demonstrate that intertidal elevation strongly affects the community characteristics and population dynamics of B. sexangula. This study provides critical baseline data for the protection and optimized management of mangrove ecosystems, particularly in regions experiencing dynamic intertidal changes.
-
花,作为植物的繁殖器官,主要由花瓣、雄蕊、雌蕊、花托等组织构成。不同种类植物的花在结构上存在差异,这些组织和结构的独特性和完整性对于植物实现传粉与繁殖成功至关重要。研究发现,大约90%的开花植物依赖动物传粉,随着全球传粉者多样性的下降[1-5],如何保证植物能够获得充足的传粉服务已成为一个重要的问题[6]。近年来,关于植物、花微生物与传粉者之间相互作用的研究日益受到关注,研究认为花微生物在维持植物与传粉者相互作用关系中起到了重要作用[7]。花为微生物提供丰富的营养物质和适宜生长的环境,有利于微生物的定殖[8]。定殖于花上的微生物通过干扰花粉萌发、花粉管生长以及果实发育等过程,直接影响植物的有性生殖过程[9-11];也可以通过改变花信号,影响传粉者行为及组成,间接影响植物传粉成功[12-15]。为研究花微生物对植物传粉与繁殖成效的影响,首先需要了解花微生物的组成及群落构建过程。花表面承载着丰富的微生物群落,包括细菌、真菌和病毒等[16-21]。根据微生物的来源,可以将其分为非生物来源与生物来源。非生物来源的微生物主要指来自空气、土壤及雨水等自然环境因素的微生物[22],而生物来源的微生物则主要包括传粉者携带并移入花中的微生物以及植物自身内源性的微生物。因此,花表面微生物的群落组成受到植物自身特性、环境因素及传粉者访问等因素的共同影响[23, 24]。研究植物与传粉者等因素对花微生物群落构建的影响,有助于深入理解植物、花微生物与传粉者之间的三方互作关系。
在花微生物的相关研究中,研究者发现花蜜微生物可以影响植物的传粉与繁殖成功,花蜜微生物多样性及功能的研究已成为该领域热点之一[9-15],而对花药微生物多样性及功能的研究相对较少。花药在植物繁殖过程中发挥着重要作用,其表面也通常较为粗糙,便于微生物的定殖[25]。花药同时也是一些植物病原体易感染的部位,容易引起植物居群内产生严重的与花粉相关的病害[26, 27],例如微球黑粉菌(Microbotryum)可以在石竹科植物的花药上快速繁殖,而传粉者作为花粉与病原菌的载体,可以将病原菌扩散到新的植株上,最终导致整个居群的感染[28-31]。Curran等[32]的研究表明,花药感染黑粉菌后,酢浆草属(Oxalis)植物的结籽数显著下降。同时,一些与花粉或柱头相关的微生物可能进入宿主的繁殖过程,作为种子内生菌传播并感染下一代[33]。因此,理解花药微生物的组成及群落构建模式对于提高植物的繁殖适合度具有潜在意义。作为冬季开花的果树,枇杷(Eriobotrya japonica (Thunb.) Lindl.)为冬季传粉者提供了重要的蜜粉资源,同时枇杷依赖传粉者传递花粉以实现繁殖成功[34]。枇杷单花约含20枚花药,可以释放大量花粉。研究表明,枇杷在花期和果期容易感染多种真菌与细菌病害[35, 36],造成枇杷果实产量与品质的下降,然而目前相关研究仍相对缺乏。
在以往的研究中,研究者主要使用传统的平板培养技术统计花微生物的发生率和密度[16, 37, 38],但这种方法存在一定的限局性,如无法培养出对环境或营养条件有特殊需求的微生物。随着科技的发展,高通量测序技术在环境科学、食品工业等领域得到广泛应用,可以帮助研究者更准确地解析样本中微生物的种类及相对含量,并全面分析微生物群落的多样性以及分布规律[39, 40]。本文拟通过高通量测序技术探究枇杷花药中真菌与细菌的群落组成模式,以期为该物种繁殖过程中多物种的互作研究提供参考资料。
1. 材料与方法
1.1 实验材料
枇杷属于蔷薇科枇杷属(Eriobotrya),为常绿小乔木,适宜在温暖环境中生长,在四川、贵州、湖北等地被广泛种植。枇杷单个花序上可以生长约100~200朵花,每朵花都具有蜜腺、5个花柱以及约20个花药。本文研究地点位于湖北省武汉市武汉大学校内(30°32′06″N,114°21′19″E,海拔82.5 m),该区域人工种植枇杷植株的花期从11月持续至次年1月,果期大约在5月左右。枇杷的单花期最长可达7 d,柱头在开花第1天内即具有可授性,而花药在开花第2天逐渐纵裂并释放花粉。枇杷花的访问者包括中华蜜蜂(Apis cerana),墨胸胡蜂(Vespa velutina)以及红头丽蝇(Calliphora vicina)等。在较为温暖的11月份,传粉昆虫的访花高峰期为9:00-16:00,并且受到温度的影响,每朵枇杷单花在10 min内接受传粉昆虫的访问次数范围为0.1~0.4次[34]。
1.2 实验方法
1.2.1 花药的收集
为研究传粉者对枇杷花药微生物群落组成的影响,设置套袋组与访问组。套袋组使用尼龙网纱袋对花序进行套袋处理,以此排除传粉者的访问;访问组则不进行套袋处理,允许传粉者自由访问枇杷单花。在正式实验开始前,移除实验组内已开放的枇杷花,以保证两组中花期的一致性。于2021年11月,在6个枇杷植株上随机选取60个花序分别作为对照组(30个花序)与访问组(30个花序),每个花序中至少含有2~3朵枇杷单花。优先选取单花期在第3天的枇杷花作为实验样本(花药已全部开裂),此阶段访问组的枇杷花在天气晴朗时更可能接收到传粉者访问。使用消毒后的剪刀收集套袋组与访问组枇杷单花,分别放入网纱袋中,并就地取样。随后使用消毒后的镊子挑取花药放入EP管中,将20朵枇杷单花的全部花药作为一个样本,套袋组与访问组各收集4个样本。为了消除天气、温度等环境因素对花药微生物的组成产生的潜在影响,在一周内对花药进行多次取样,减少随机误差,提高数据的准确性。将收集的花药样本放置于冰盒中,随即带回实验室置于−80 ℃的冰箱中冷冻保存,防止微生物的继续增殖;最后,将花药样本寄往美吉公司进行扩增子测序(16S与ITS测序)。
1.2.2 数据分析
为比较传粉者访问对花药微生物Alpha多样性的影响,使用Wilcoxon秩和检验分别比较套袋组与访问组中细菌与真菌的Alpha多样性,并使用BH进行多重检验校正。此外,为了评估传粉者访问对花药微生物组成的影响,分别计算组内及组间的Bray-Curtis、Unweighted-UniFrac和Weighted-UniFrac距离,并使用置换多元方差分析(PERMANOVA)进行差异性检验。最后,通过非度量多维尺度(NMDS)对套袋组与访问组细菌和真菌组成的差异进行可视化呈现。
2. 结果与分析
2.1 花药微生物稀释曲线
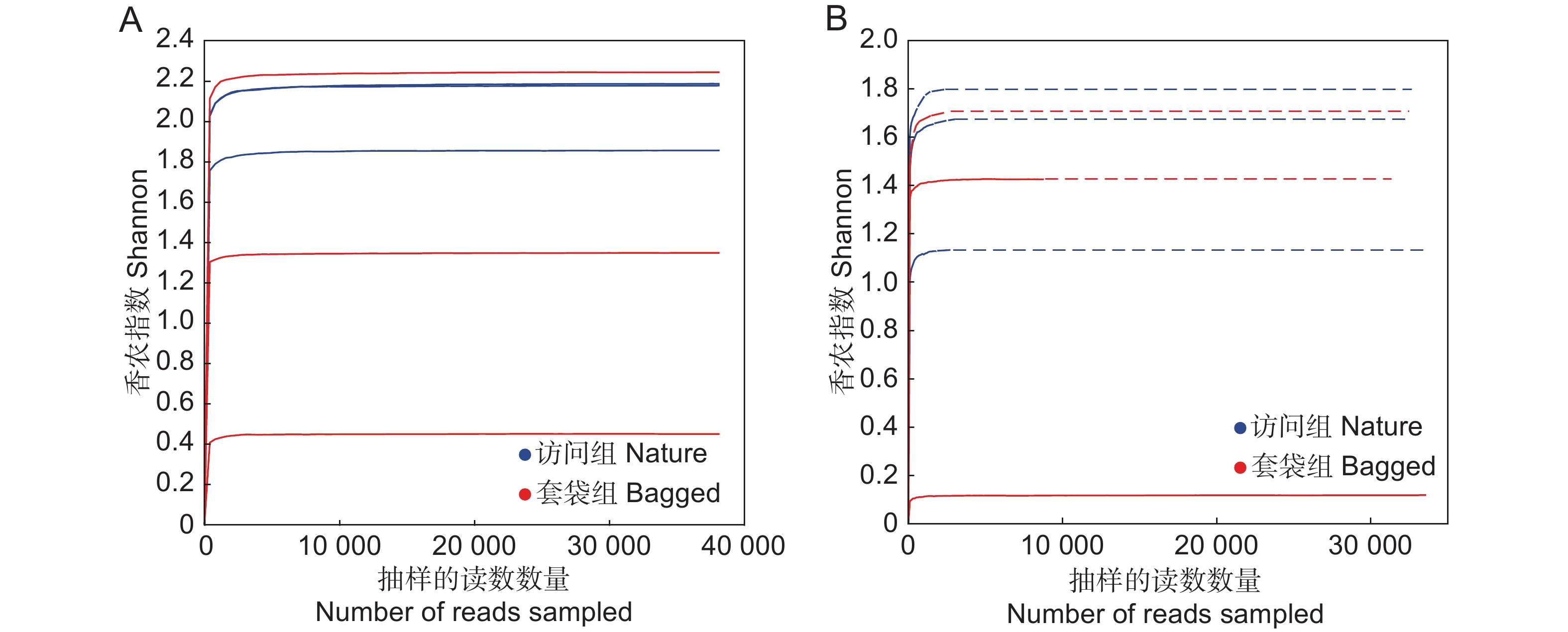
基于Shannon指数绘制套袋组与访问组中花药真菌与细菌的稀释曲线。结果显示,随着测序深度的加深,两组处理中花药真菌与细菌稀释曲线趋于平缓,并最终达到饱和状态(图1)。表明在该测序深度下,绝大多数的真菌与细菌均已被检测到。因此,测序结果可以反映枇杷花药在接受传粉者访问前后的微生物的组成及多样性。
![]() 图 1 枇杷花药微生物群落稀释曲线基于香农指数(Shannon)展示枇杷套袋组(红色)与访问组(蓝色)花药真菌(A)与细菌(B)群落的稀释曲线。Figure 1. Rarefaction curves of microbial community in anthers of Eriobotrya japonica based on Shannon indexRarefaction curves based on Shannon index, showing fungal (A) and bacterial (B) diversity in bagged (red) and nature (blue) groups.
图 1 枇杷花药微生物群落稀释曲线基于香农指数(Shannon)展示枇杷套袋组(红色)与访问组(蓝色)花药真菌(A)与细菌(B)群落的稀释曲线。Figure 1. Rarefaction curves of microbial community in anthers of Eriobotrya japonica based on Shannon indexRarefaction curves based on Shannon index, showing fungal (A) and bacterial (B) diversity in bagged (red) and nature (blue) groups.2.2 花药微生物群落组成
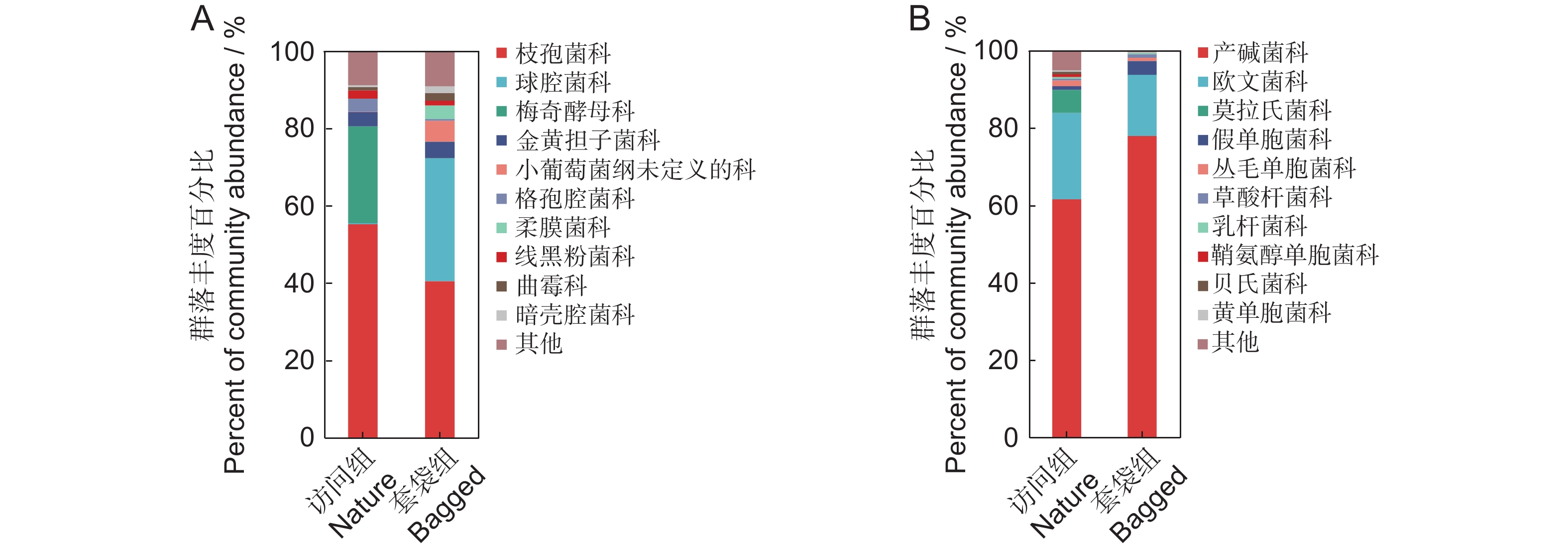
根据丰度对花药微生物进行排序,并在科的水平上列举排名前10的微生物物种,其余物种合并为“Others”(图2)。套袋组中优势细菌为产碱杆菌科、欧文菌科以及假单胞菌科,平均相对丰度分别为77.96%、15.80%和3.59%,合计占细菌总丰度的97.35%;访问组中优势细菌为产碱杆菌科、欧文菌科以及莫拉氏菌科,平均相对丰度分别为61.56%、22.36%和5.96%,合计占细菌总丰度的89.88%。套袋组中优势真菌为枝孢菌科、球腔菌科以及微球黑粉菌目下未定义的科,平均相对丰度分别为40.49%、31.74%和5.40%,占真菌总丰度的77.63%;访问组中优势真菌为枝孢菌科、梅奇酵母科以及短梗霉科,平均相对丰度分别为55.20%、25.01%和3.74%,占真菌总丰度的83.7%。对于花药细菌而言,两个实验组中都以产碱杆菌科与欧文菌科作为主要的两种细菌科,而在真菌方面,两个实验组中枝孢菌科都具有较高的占比。两个实验组中最优势的微生物均为同一科的微生物,因此花药中具有相对稳定的微生物群落。
![]() 图 2 枇杷花药微生物群落结构(科水平)A:以ITS2测序表征的平均丰度>1%的套袋组与访问组真菌科组成;B:以16S测序表征的平均丰度>1%的套袋组与访问组细菌科组成。图中仅展示各实验组中丰度占比前10的微生物,其余物种合并为“其他”。Figure 2. Structure of microbial community in anthers of Eriobotrya japonica (family level)A: Composition of fungal families with an average abundance greater than 1%, characterized by ITS2 sequencing, in nature and bagged groups; B: Composition of bacterial families with an average abundance greater than 1%, characterized by 16S sequencing, in nature and bagged groups. Only the top 10 most abundant microbes in each experimental group are shown in the figure, with all other species combined under “Others”.
图 2 枇杷花药微生物群落结构(科水平)A:以ITS2测序表征的平均丰度>1%的套袋组与访问组真菌科组成;B:以16S测序表征的平均丰度>1%的套袋组与访问组细菌科组成。图中仅展示各实验组中丰度占比前10的微生物,其余物种合并为“其他”。Figure 2. Structure of microbial community in anthers of Eriobotrya japonica (family level)A: Composition of fungal families with an average abundance greater than 1%, characterized by ITS2 sequencing, in nature and bagged groups; B: Composition of bacterial families with an average abundance greater than 1%, characterized by 16S sequencing, in nature and bagged groups. Only the top 10 most abundant microbes in each experimental group are shown in the figure, with all other species combined under “Others”.2.3 花药微生物多样性差异
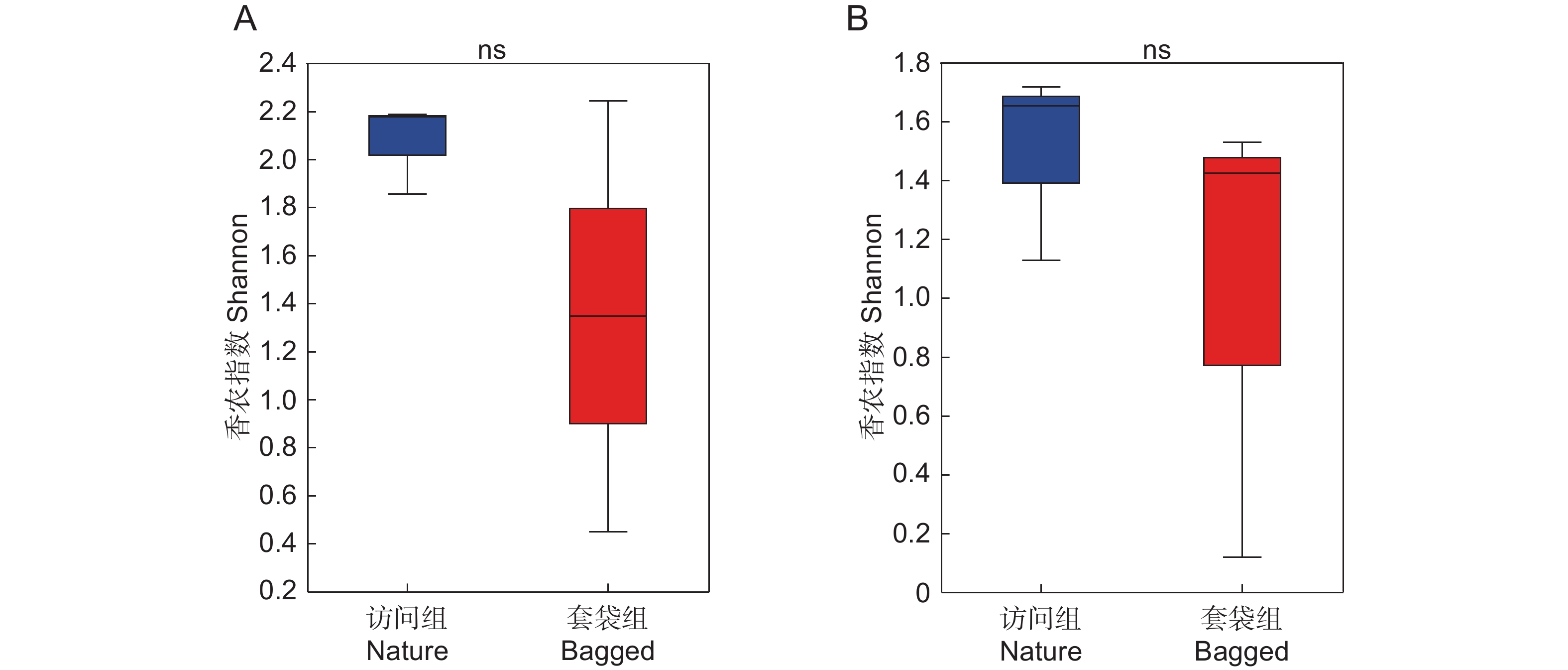
基于Shannon指数计算套袋组与访问组花药中细菌与真菌群落的Alpha多样性,并比较两组之间的差异。结果显示,与套袋组相比,访问组中细菌与真菌的Alpha多样性均呈增加趋势,但在统计学上并未达到显著性水平(Wilcoxon秩和检验,BH校正,P细菌=0.993 8,P真菌=0.662 5,图3)。此外,本文计算了Chao以及Coverage指数,从微生物群落的丰富度和覆盖度角度分析套袋处理对细菌和真菌群落Alpha多样性的影响。基于Chao与Coverage指数对比两组Alpha多样性差异,发现套袋处理对微生物群落Alpha多样性没有显著影响,因此并未在本文中呈现详细数据。
![]() 图 3 枇杷花药微生物Alpha多样性分析(Shannon指数)基于香农指数展示枇杷访问组(蓝色)与套袋组(红色)花药真菌(A)与细菌(B)的Alpha多样性。利用Wilcoxon秩和检验比较两个实验组中真菌与细菌Alpha多样性的差异,箱线图显示了中值和四分位距,ns表示两个实验组间不存在显著差异。Figure 3. Alpha diversity analysis (Shannon index) of microbial community in anthers of Eriobotrya japonicaAlpha diversity of fungal (A) and bacterial (B) communities in anthers of Eriobotrya japonica in nature (blue) and bagged (red) groups is presented based on Shannon index. Differences in fungal and bacterial alpha diversity between two experimental groups were assessed using Wilcoxon rank-sum test. Boxplot illustrates median and interquartile range, with “ns” signifying no significant difference.
图 3 枇杷花药微生物Alpha多样性分析(Shannon指数)基于香农指数展示枇杷访问组(蓝色)与套袋组(红色)花药真菌(A)与细菌(B)的Alpha多样性。利用Wilcoxon秩和检验比较两个实验组中真菌与细菌Alpha多样性的差异,箱线图显示了中值和四分位距,ns表示两个实验组间不存在显著差异。Figure 3. Alpha diversity analysis (Shannon index) of microbial community in anthers of Eriobotrya japonicaAlpha diversity of fungal (A) and bacterial (B) communities in anthers of Eriobotrya japonica in nature (blue) and bagged (red) groups is presented based on Shannon index. Differences in fungal and bacterial alpha diversity between two experimental groups were assessed using Wilcoxon rank-sum test. Boxplot illustrates median and interquartile range, with “ns” signifying no significant difference.2.4 花药微生物组成差异
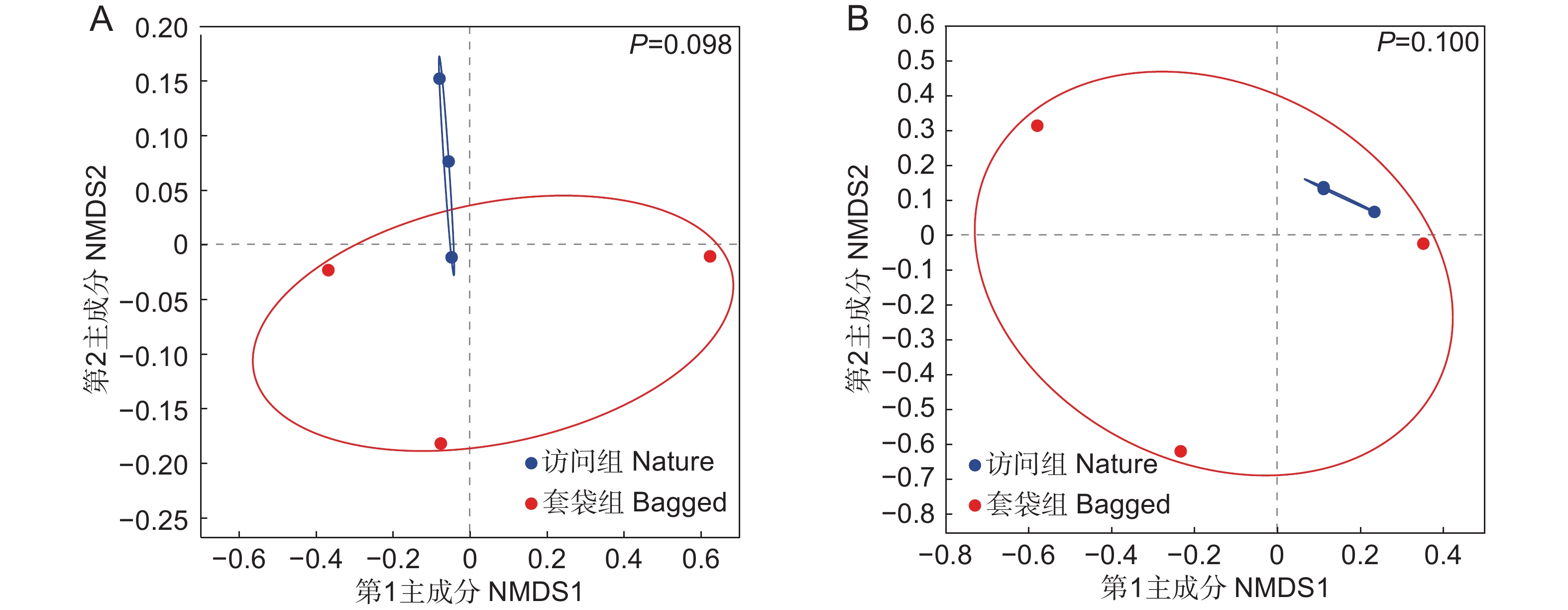
基于Bray-Curtis距离,对套袋组和访问组枇杷花药微生物群落进行了NMDS排序。结果表明,两个实验组中细菌与真菌群落的组成没有显著差异(图4),即套袋处理不会改变花药微生物的群落组成。Bray-Curtis距离算法仅考虑每个样本中微生物的相对丰度,Unweighted-UniFrac距离算法则只关注微生物分类单元的进化关系,Weighted-UniFrac距离算法综合考虑了微生物的相对丰度及分类单元的进化关系。在使用不同的距离算法进一步分析套袋处理对微生物分类单元进化关系的影响时,结果与Bray-Curtis距离算法相一致(表1)。因此,花药微生物群落组成具有较高的稳定性,不受套袋处理的影响。
![]() 图 4 枇杷花药微生物群落的NMDS分析基于Bray-Curtis距离对枇杷花药微生物群落进行非度量多维尺度(NMDS)排序,对真菌(A)和细菌(B)进行了单独排序。蓝色实心圆圈代表访问组样本,红色实心圆圈代表套袋组样本。绘制分组椭圆展示样本点之间的分布情况。基于Bray-Curtis距离算法,利用PERMANOVA比较套袋组与访问组真菌与细菌群落组成,P<0.05表示具有显著差异。Figure 4. Non-metric multidimensional scaling (NMDS) of microbial community in anthers of Eriobotrya japonicaNon-metric multidimensional scaling (NMDS) ordination of microbial communities in anthers of Eriobotrya japonica based on Bray-Curtis distance was performed, with separate analyses for fungi (A) and bacteria (B). Blue solid circles represent samples from nature group, red solid circles represent samples from bagged group. Group ellipses are drawn to present distribution of sample points. Based on Bray-Curtis distance algorithm, PERMANOVA was used to compare fungal and bacterial community compositions between bagged and nature groups, with P<0.05 indicating significant differences.表 1 使用不同距离算法的PERMANOVA分析汇总Table 1. Summary of permutation multivariate analysis of variance (PERMANOVA) using different distance algorithms
图 4 枇杷花药微生物群落的NMDS分析基于Bray-Curtis距离对枇杷花药微生物群落进行非度量多维尺度(NMDS)排序,对真菌(A)和细菌(B)进行了单独排序。蓝色实心圆圈代表访问组样本,红色实心圆圈代表套袋组样本。绘制分组椭圆展示样本点之间的分布情况。基于Bray-Curtis距离算法,利用PERMANOVA比较套袋组与访问组真菌与细菌群落组成,P<0.05表示具有显著差异。Figure 4. Non-metric multidimensional scaling (NMDS) of microbial community in anthers of Eriobotrya japonicaNon-metric multidimensional scaling (NMDS) ordination of microbial communities in anthers of Eriobotrya japonica based on Bray-Curtis distance was performed, with separate analyses for fungi (A) and bacteria (B). Blue solid circles represent samples from nature group, red solid circles represent samples from bagged group. Group ellipses are drawn to present distribution of sample points. Based on Bray-Curtis distance algorithm, PERMANOVA was used to compare fungal and bacterial community compositions between bagged and nature groups, with P<0.05 indicating significant differences.表 1 使用不同距离算法的PERMANOVA分析汇总Table 1. Summary of permutation multivariate analysis of variance (PERMANOVA) using different distance algorithmsGroup df Bray-Curtis Unweighted-UniFrac Weighted-UniFrac F R2 P F R2 P F R2 P 真菌 1 1.423 0.262 0.098 1.387 0.257 0.100 1.233 0.236 0.200 细菌 1 2.167 0.351 0.100 0.991 0.198 0.500 0.473 0.106 0.900 3. 讨论
花粉的“健康”直接影响植物的繁殖成功,因此维护花药内部环境的稳定性至关重要。研究表明,当花药感染某些微生物后,会对植物的生殖适合度产生消极影响[32],同时传粉者访问会改变花微生物的群落组成[41]。本文通过高通量测序技术探究了传粉者访问前(套袋组)与传粉者访问后(访问组),人工种植的枇杷花药微生物的组成及其变化。结果表明,两组花药微生物的Alpha多样性不存在显著差异,并且最优势的花药微生物属于同一个科。在细菌方面,两组均以产碱杆菌科和欧文菌科为主要细菌;而在真菌方面,枝孢菌科在两组中具有较高的占比。
本文利用不同距离算法比较了套袋组与访问组微生物群落组成,发现花药微生物群落组成具有较强的稳定性,并且传粉者不是驱动花药微生物群落构建的主要因素。研究普遍认为,传粉者的访问会改变花微生物群落的组成[21, 42, 43],Pozo等[41]在6个不同海拔研究了同一植物的酵母发生率与丰度,发现传粉者的组成与活动对花微生物群落的构建具有显著影响。然而,Rebolleda Gómez等[23]研究了花部位、传粉者及不同居群对多斑沟酸浆(Mimulus guttatus Fisch. ex DC.)花微生物群落组成的影响,发现不同花部位和不同居群之间的微生物群落组成具有显著差异,而传粉者的访问对微生物组成并没有显著影响。因此,本研究认为花的不同部位存在过滤机制,并且是影响微生物群落构建的主要因素。研究者已经发现叶片及花蜜对微生物的定殖存在过滤机制,而对花药微生物是否存在过滤机制的研究相对较少。Vokou等[44]比较了空气细菌与9种植物叶片附生细菌组成的相似性,发现只有少数空气细菌适应叶片环境并定殖。花蜜则通过高渗透压、高糖浓度等生理特性,限制了微生物的定殖[38, 45]。Junker等[46]利用高通量测序技术对比分析了多型铁心木(Metrosideros polymorpha Gaudich.)叶片、花药、花蜜以及柱头细菌群落的组成,发现与海拔,地理距离等因素相比,不同部位潜在的过滤机制是影响微生物群落组成的主要因素。Ambika Manirajan等[33]比较了4种植物花粉微生物的组成,结果显示不同花粉的微生物组成具有显著差异,并认为不同植物花粉独特的营养水平与结构特征,导致了这种差异。Kosenko等[47]利用扫描电镜解析了34种植物花粉的超微结构,发现不同植物(属或种)花粉粒外层的形状与结构具有显著差异,侧面印证了Ambika Manirajan等[33]的猜测。因此,花药存在过滤机制影响其表面微生物的群落构建。此外,有研究表明花蜜与花柱微生物存在优先效应,即优先定殖的微生物抢占营养及空间资源,进而限制后续微生物定殖[13, 48-50]。由于优先效应的存在,花蜜与花柱微生物物种多样性较低,微生物组成由单一的优势酵母与细菌为主导[45, 51],这与本文花药的研究结果一致。在本研究中,尽管传粉者的访问会改变某些微生物的相对含量(引入新的微生物),但整体的微生物群落组成并未发生显著变化,因此我们认为花药存在的过滤机制及花药微生物存在的优先效应可能是枇杷花药微生物群落构建的主要因素。
由于实验条件限制,本研究仅使用扩增子测序技术探索了传粉者访问前后,枇杷花药中微生物的多样性,而未通过传统的培养技术获得花药中优势微生物,并进一步探索微生物的功能。研究表明,花蜜微生物会通过直接或间接的方式影响植物的传粉与繁殖成效[9-15]。例如微生物通过改变视觉、嗅觉或味觉信号来影响传粉者的访问行为,进而影响植物的繁殖成效[15, 52-55],或是直接干扰柱头花粉萌发影响植物的繁殖过程[13]。与花蜜微生物功能类似,花药微生物可能存在潜在的功能,但对于花药上真菌和细菌群落如何影响传粉后植物的繁殖成功还未见报道。
此外,枇杷花具有多种传粉者,我们未探讨这些传粉者携带的微生物是否具有差异,携带的微生物是否会定殖在花药上并产生新的花信号,进而对枇杷的传粉与繁殖成效产生不同的影响。Morris等[56]比较了柳叶菜属植物(Epilobium canum (Greene) P. H. Raven.)在接受蜂鸟访问以及木蜂盗取花蜜后,花蜜微生物群落组成的差异,发现花蜜在接受蜂鸟与木蜂访问后,微生物丰度显著增加,并且在木蜂盗取花蜜后,花蜜微生物丰度是蜂鸟访问后花蜜微生物丰度的10倍。此外,与蜂鸟访问后的花蜜相比,木蜂实施盗取行为后的花蜜中,醋酸菌科与梅奇酵母科成为新的优势微生物。作者因此认为传粉者类型也是驱动花蜜微生物群落构建的主要因素。
花药微生物群落组成同样受到植物种类的影响[33, 46]。Ambika Manirajan等[33]利用高通量测序技术检测了垂枝桦(Betula pendula Roth.)、黑麦(Secale cereale L.)、欧洲油菜(Brassica napus L.)、秋水仙(Colchicum autumnale L.)花粉细菌的群落组成,发现垂枝桦、欧洲油菜、秋水仙的花粉微生物以肠杆菌科为主,而微球菌科和诺卡氏菌科是黑麦花粉中的优势微生物。Junker等[46]测定了多型铁心木花药微生物的组成,同样发现肠杆菌科是多型铁心木花药的优势细菌。因此,肠杆菌科微生物可能是花药中较为常见的一类细菌。在本文中,枇杷花药微生物中以产碱杆菌、欧文菌科为优势细菌,这与Ambika Manirajan[33]、Junker等[46]的研究结果具有差异。我们认为本文的研究对象枇杷,其传粉者与传粉类型(依靠中华蜜蜂、墨胸胡蜂等进行虫媒传粉)与上述5种植物的传粉者与传粉类型存在差异(风媒传粉或依靠西部黄胡蜂等传粉者进行虫媒传粉),进而导致优势微生物种类存在差异。此外,本文是在武汉大学校园内对枇杷花药微生物群落进行的相关研究,该区域枇杷定期接受人工管理,因此其微生物组成也可能受到人类行为的影响。目前也有研究探讨了果树花蜜微生物的群落组成及其对植物繁殖适合度的影响[39, 40, 57]。将本文花药微生物的群落组成与上述文献对比,发现在科的水平上,枇杷花药优势微生物与苹果(Malus domestica Borkh.)、西洋梨(Pyrus communis L.)、高丛越橘(Vaccinium corymbosum L.)、番茄(Solanum lycopersicum L.)的花蜜优势微生物不属于同一科,该结果可能是由于不同花部位的过滤机制导致;而梨(Pyrus)、蓝莓(Vaccinium uliginosum L.)、番茄中花蜜优势微生物也不相同,该结果可能是由于植物种类不同导致。对于花药微生物群落组成机制,以及微生物潜在功能的探讨,有助于深入理解植物、花微生物与传粉者之间的三方互作关系,并为果树繁殖过程中微生物的多样性及生态功能的研究提供参考资料。
-
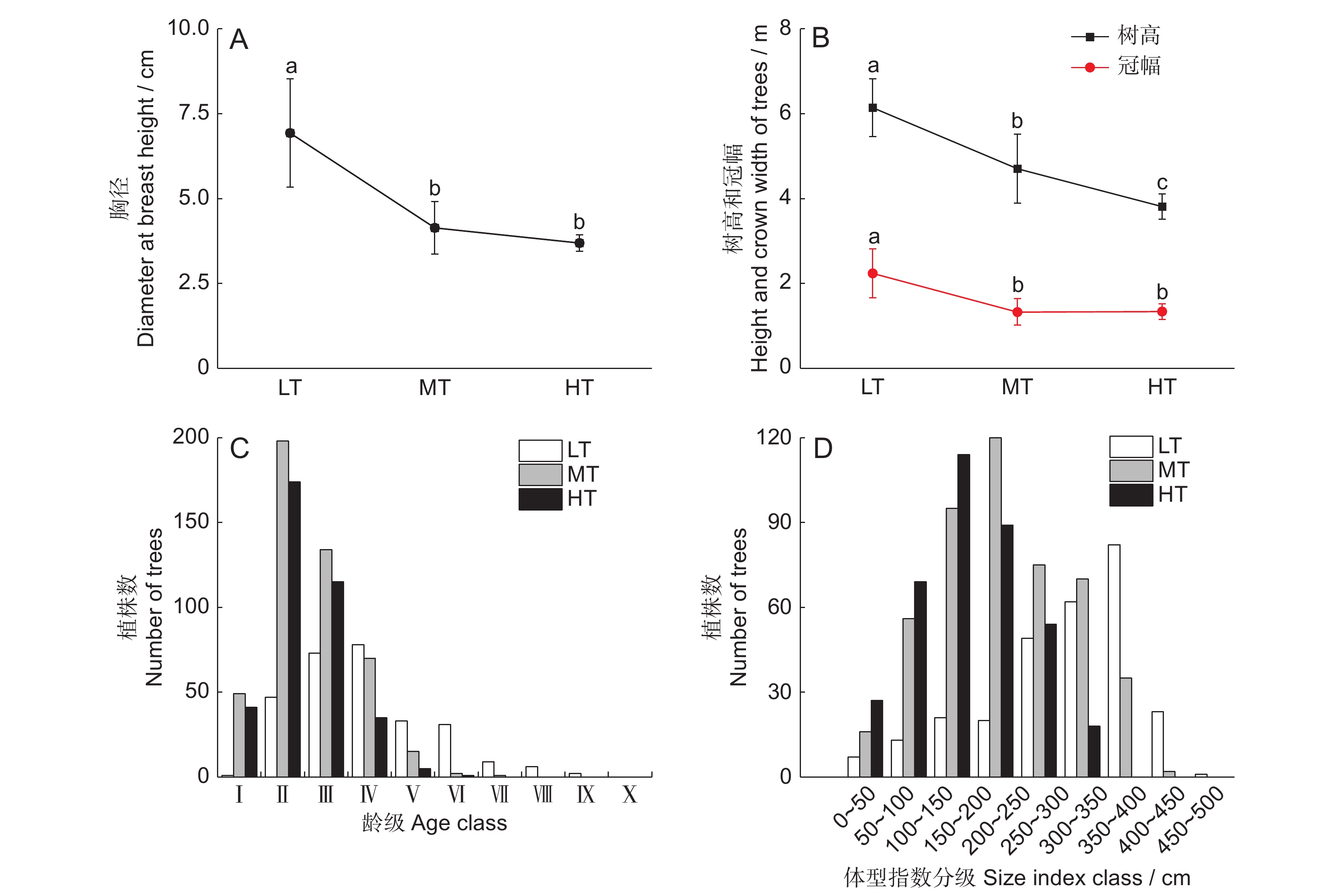
![]()
图 1 不同潮位生境海莲群落的特征及林分密度
不同小写字母表示差异显著(P<0.05)。LT:低潮位;MT:中潮位;HT:高潮位。下同。
Figure 1. Community characteristics and stand density of Bruguiera sexangula at different intertidal elevations
Different lowercase letters indicate significant differences (P<0.05). LT: Low intertidal elevation; MT: Middle intertidal elevation; HT: High intertidal elevation. Same below.
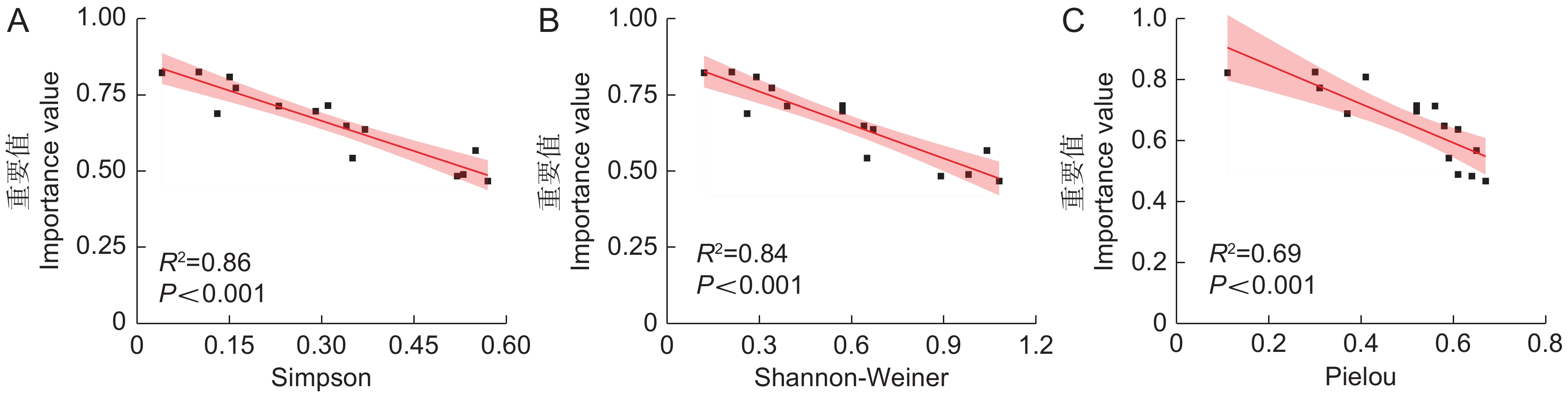
![]()
图 2 海莲重要值与群落多样性指数间的相关性分析
Figure 2. Correlation analysis between importance value and community diversity index in Bruguiera sexangula
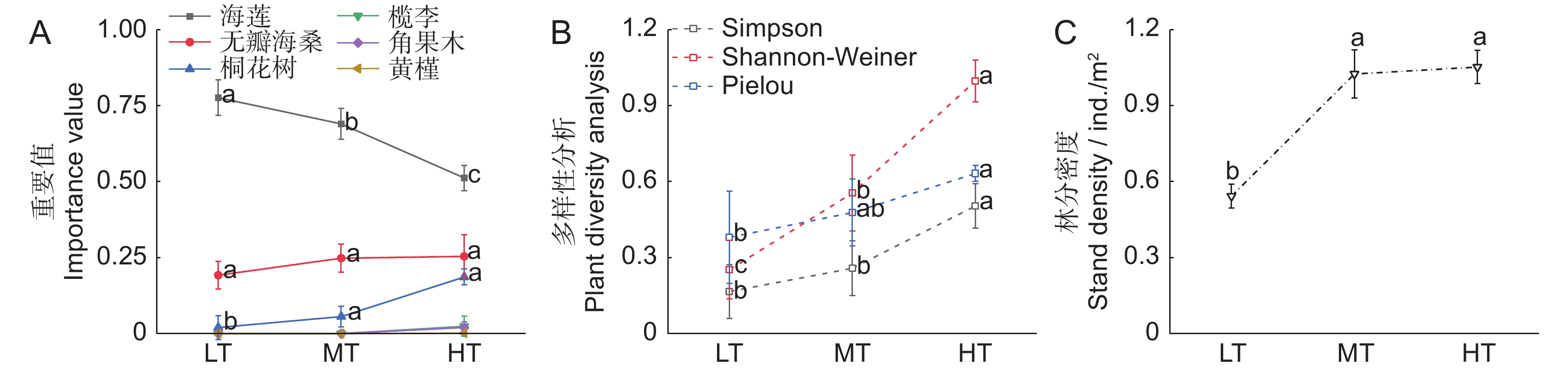
![]()
图 3 不同潮位生境海莲种群的基本特征
Figure 3. Population characteristics of Bruguiera sexangula at different intertidal elevations
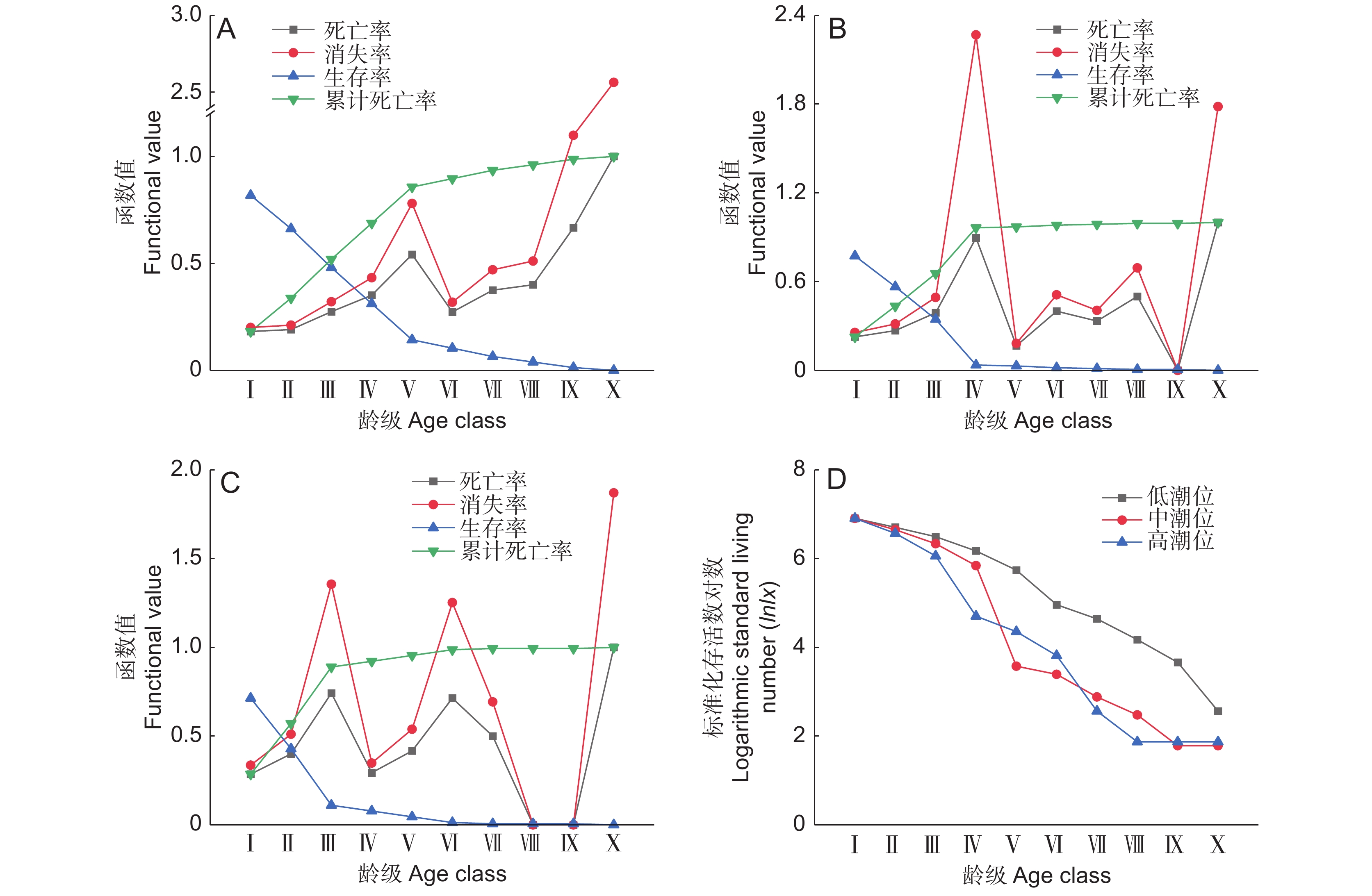
![]()
图 4 不同潮位生境海莲种群的生存分析
A:低潮位;B:中潮位;C:高潮位;D:不同潮位生境下海莲种群的存活曲线。
Figure 4. Population dynamics of Bruguiera sexangula at different intertidal elevations
A: Low intertidal elevation; B: Middle intertidal elevation; C: High intertidal elevation; D: Survival curve for Bruguiera sexangula population at different intertidal elevations.
表 1 不同潮位下海莲群落内的植物种类和植株数量
Table 1 Plant species and numbers in Bruguiera sexangula communities at different intertidal levels
物种Species 株数Number 低潮位
Low intertidal
elevation中潮位
Middle intertidal
elevation高潮位
High intertidal
elevation海莲Bruguiera sexangula (Lour.) Poiret 280 469 371 无瓣海桑Sonneratia apetala Buchanan-Hamilton 17 34 28 桐花树Aegiceras corniculatum (L.) Blanco 10 45 149 榄李Lumnitzera racemosa Willdenow 0 0 13 角果木Ceriops tagal (Perr.) CB Robinson 0 0 15 黄槿Talipariti tiliaceum (L.) Fryxell 0 0 1  下载: 导出CSV
下载: 导出CSV
表 2 不同潮位生境海莲种群的分布格局
Table 2 Distribution pattern of Bruguiera sexangula populations at different intertidal elevations
潮位
Intertidal zonem* P K C I Ca 分布格局
Distribution patternLT 59.72 1.07 15.05 4.72 3.72 0.07 C MT 96.90 1.03 32.46 3.90 2.90 0.03 C HT 76.15 1.03 34.35 3.15 2.15 0.03 C 注:C,聚集分布;m*,平均拥挤度;P,聚块性指数;K,负二项式指数;C,扩散系数;I,丛生指数;Ca,Cassie指标;LT,低潮位;MT,中潮位;HT,高潮位。 Notes: C, Clumped distribution; m*, Average congestion; P, Agglomerative index; K, Negative binomial index; C, Diffusion coefficient; I, Overgrowth index; Ca, Cassie’s Indicator; LT, Low intertidal elevation; MT, Middle intertidal elevation; HT, High intertidal elevation.
下载: 导出CSV
表 3 不同潮位生境海莲种群的静态生命表
Table 3 Static life table parameters of Bruguiera sexangula at different intertidal elevations
潮位
Intertidal zone龄级
Age classAx ax lx lnlx dx qx Lx Tx ex Kx Sx LT Ⅰ 1 77 1 000.00 6.91 181.82 0.18 909.09 3136.36 3.45 0.20 0.82 Ⅱ 47 63 818.18 6.71 155.84 0.19 740.26 2227.27 3.01 0.21 0.81 Ⅲ 73 51 662.34 6.50 181.82 0.27 571.43 1487.01 2.60 0.32 0.73 Ⅳ 78 37 480.52 6.17 168.83 0.35 396.10 915.58 2.31 0.43 0.65 Ⅴ 33 24 311.69 5.74 168.83 0.54 227.27 519.48 2.29 0.78 0.46 Ⅵ 31 11 142.86 4.96 38.96 0.27 123.38 292.21 2.37 0.32 0.73 Ⅶ 9 8 103.90 4.64 38.96 0.38 84.42 168.83 2.00 0.47 0.63 Ⅷ 6 5 64.94 4.17 25.97 0.40 51.95 84.42 1.63 0.51 0.60 Ⅸ 2 3 38.96 3.66 25.97 0.67 25.97 32.47 1.25 1.10 0.33 Ⅹ 0 1 12.99 2.56 12.99 1.00 6.49 6.49 1.00 2.56 0.00 MT Ⅰ 49 168 1 000.00 6.91 226.19 0.23 886.90 2 291.67 2.58 0.26 0.77 Ⅱ 198 130 773.81 6.65 208.33 0.27 669.64 1 404.76 2.10 0.31 0.73 Ⅲ 134 95 565.48 6.34 220.24 0.39 455.36 735.12 1.61 0.49 0.61 Ⅳ 70 58 345.24 5.84 309.52 0.90 190.48 279.76 1.47 2.27 0.10 Ⅴ 15 6 35.71 3.58 5.95 0.17 32.74 89.29 2.73 0.18 0.83 Ⅵ 2 5 29.76 3.39 11.90 0.40 23.81 56.55 2.38 0.51 0.60 Ⅶ 1 3 17.86 2.88 5.95 0.33 14.88 32.74 2.20 0.41 0.67 Ⅷ 0 2 11.90 2.48 5.95 0.50 8.93 17.86 2.00 0.69 0.50 Ⅸ 0 1 5.95 1.78 0.00 0.00 5.95 8.93 1.50 0.00 1.00 Ⅹ 0 1 5.95 1.78 5.95 1.00 2.98 2.98 1.00 1.78 0.00 HT Ⅰ 41 154 1 000.00 6.91 285.71 0.29 857.14 1 909.09 2.23 0.34 0.71 Ⅱ 174 110 714.29 6.57 285.71 0.40 571.43 1051.95 1.84 0.51 0.60 Ⅲ 115 66 428.57 6.06 318.18 0.74 269.48 480.52 1.78 1.36 0.26 Ⅳ 35 17 110.39 4.70 32.47 0.29 94.16 211.04 2.24 0.35 0.71 Ⅴ 5 12 77.92 4.36 32.47 0.42 61.69 116.88 1.89 0.54 0.58 Ⅵ 1 7 45.45 3.82 32.47 0.71 29.22 55.19 1.89 1.25 0.29 Ⅶ 0 2 12.99 2.56 6.49 0.50 9.74 25.97 2.67 0.69 0.50 Ⅷ 0 1 6.49 1.87 0.00 0.00 6.49 16.23 2.50 0.00 1.00 Ⅸ 0 1 6.49 1.87 0.00 0.00 6.49 6.49 1.00 0.00 1.00 Ⅹ 0 1 6.49 1.87 6.49 1.00 3.25 3.25 1.00 1.87 0.00 注:Ax,x龄级内存活的个体数;ax,匀滑修正后x龄级内存活的个体数;lx,x龄级的标准化存活个体数;lnlx,lx的自然对数;dx,x龄级到x+1龄级间的标准化死亡数;qx,x龄级到x+1龄级间的死亡率;Lx,x龄级到x+1龄级间平均存活个体数;Tx,x龄级到超过x龄级的个体总数;ex,进入x龄级个体的期望寿命;Kx,消失率;Sx,存活率。 Notes: Ax, Individual number of age class x; ax, Revised data of Ax,; lx, Standardized number of surviving individuals of age class x; lnlx, Natural logarithm of lx; dx, Standardized number of mortality from age class x to age class x+1; qx, Mortality rate from age class x to age class x+1; Lx, Number of surviving individuals from age class x to age class x+1; Tx, Total individuals of age class x and age class older than x; ex, Life expectancy of individuals entering age class x; Kx, Vanish rate; Sx, Survival rate.
下载: 导出CSV
-
[1] De Deyn GB,Raaijmakers CE,van der Putten WH. Plant community development is affected by nutrients and soil biota[J]. J Ecol,2004,92(5):824−834. doi: 10.1111/j.0022-0477.2004.00924.x
[2] 方精云,王襄平,沈泽昊,唐志尧,贺金生,等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性,2009,17(6):533−548. doi: 10.3724/SP.J.1003.2009.09253 Fang JY,Wang XP,Shen ZH,Tang ZY,He JS,et al. Methods and protocols for plant community inventory[J]. Biodiversity Science,2009,17(6):533−548. doi: 10.3724/SP.J.1003.2009.09253
[3] Simard M,Fatoyinbo L,Smetanka C,Rivera-Monroy VH,Castañeda-Moya E,et al. Mangrove canopy height globally related to precipitation,temperature and cyclone frequency[J]. Nat Geosci,2019,12(1):40−45. doi: 10.1038/s41561-018-0279-1
[4] He BY,Lai TH,Fan HQ,Wang WQ,Zheng HL. Comparison of flooding-tolerance in four mangrove species in a diurnal tidal zone in the Beibu Gulf[J]. Estuar Coast Shelf S,2007,74(1-2):254−262. doi: 10.1016/j.ecss.2007.04.018
[5] Suyadi N,Gao J,Lundquist CJ,Schwendenmann L. Land-based and climatic stressors of mangrove cover change in the Auckland Region,New Zealand[J]. Aquat Conserv,2019,29(9):1466−1483. doi: 10.1002/aqc.3146
[6] Li LF,Liu W,Ai JW,Cai SJ,Dong JW. Predicting mangrove distributions in the Beibu Gulf,Guangxi,China,using the MaxEnt model:determining tree species selection[J]. Forests,2023,14(1):149. doi: 10.3390/f14010149
[7] Chen YP,Ye Y. Growth and physiological responses of saplings of two mangrove species to intertidal elevation[J]. Mar Ecol Prog Ser,2013,482:107−118. doi: 10.3354/meps10274
[8] Zhu P,Wang YP,Shi TT,Zhang XL,Huang GQ,Gong J. Intertidal zonation affects diversity and functional potentials of bacteria in surface sediments:a case study of the Golden Bay mangrove,China[J]. Appl Soil Ecol,2018,130:159−168. doi: 10.1016/j.apsoil.2018.06.003
[9] Komiyama A,Santiean T,Higo M,Patanaponpaiboon P,Kongsangchai J,Ogino K. Microtopography,soil hardness and survival of mangrove (Rhizophora apiculata BL.) seedlings planted in an abandoned tin-mining area[J]. Forest Ecol Manag,1996,81(1-3):243−248. doi: 10.1016/0378-1127(95)03591-5
[10] 李巧姿. 沿海滩涂生态因子对秋茄生长的影响分析[J]. 福建林业科技,2000,27(4):31−34. Li QZ. Analyzing influences of the ecological factor on the growth of Kandelia on a coastal shoal[J]. Journal of Fujian Forestry Science and Technology,2000,27(4):31−34.
[11] 郑文炳,吴蓓莉. 海滨木槿的滩涂生态适应性研究[J]. 农业开发与装备,2012(6):83−84. doi: 10.3969/j.issn.1673-9205.2012.06.058 [12] Cheng MCF,Ho KKY,Astudillo JC,Cannicci S. An updated ecological assessment of floral and faunal communities of mangrove habitats in Tolo Harbour and Channel,Hong Kong[J]. Reg Stud Mar Sci,2023,59:102807.
[13] 金川. 浙江人工红树林对关键环境因子的生态响应研究[D]. 北京:北京林业大学,2011:4−10. [14] 王佳燕,余雪标,周光益,王旭,李洪旺. 海南东寨港海莲群落林隙结构及其更新特征[J]. 热带作物学报,2008,29(1):109−113. doi: 10.3969/j.issn.1000-2561.2008.01.023 Wang JY,Yu XB,Zhou GY,Wang X,Li HW. Canopy gap structure and regeneration character of Bruguiera sexangula in Dongzhai Harbour,Hainan Island[J]. Chinese Journal of Tropical Crops,2008,29(1):109−113. doi: 10.3969/j.issn.1000-2561.2008.01.023
[15] 李茂金,叶储民,陈玉凯. 海南清澜港红树植物海莲种群结构与分布格局研究[J]. 热带林业,2022,50(2):8−14. doi: 10.3969/j.issn.1672-0938.2022.02.002 Li MJ,Ye CM,Chen YK. Study on population structure and distribution pattern of mangrove species Bruguiera sexangula in Qinglan Port of Hainan[J]. Tropical Forestry,2022,50(2):8−14. doi: 10.3969/j.issn.1672-0938.2022.02.002
[16] 郑文教,林鹏. 盐度对红树植物海莲幼苗的生长和某些生理生态特性的影响[J]. 应用生态学报,1992,3(1):9−14. doi: 10.3321/j.issn:1001-9332.1992.01.014 Zheng WJ,Lin P. Effect of salinity on the growth and some eco-physiological characteristics of mangrove Bruguiera sexangula seedlings[J]. Chinese Journal of Applied Ecology,1992,3(1):9−14. doi: 10.3321/j.issn:1001-9332.1992.01.014
[17] 王豪,杨小波,吕晓波,李东海,王重阳,戚春林. 海南岛东北和西北部自然红树林群落特征比较[J]. 海南大学学报(自然科学版),2024,42(2):146−154. Wang H,Yang XB,Lü XB,Li DH,Wang CY,Qi CL. Characteristics comparison of the natural mangrove communities in northeast and northwest Hainan[J]. Natural Science Journal of Hainan University (Natural Science),2024,42(2):146−154.
[18] Zaman MR,Rahman MS,Ahmed S,Zuidema PA. What drives carbon stocks in a mangrove forest? The role of stand structure,species diversity and functional traits[J]. Estuar Coast Shelf Sci,2023,295:108556. doi: 10.1016/j.ecss.2023.108556
[19] Ma L,Yang SC. Growth and physiological response of Kandelia obovata and Bruguiera sexangula seedlings to aluminum stress[J]. Environ Sci Pollut Res,2022,29(28):43251−43266. doi: 10.1007/s11356-021-17926-0
[20] Azad MS,Mollick AS,Ranon RJK,Khan MNI,Kamruzzaman M. Plasticity of leaf morphology of Bruguiera sexangula to salinity zones in Bangladesh’s Sundarbans[J]. J For Res,2022,33(6):1857−1866. doi: 10.1007/s11676-022-01470-9
[21] Khanbo S,U-Thoomporn S,Kongkachana W,Charoensri S,Narong N,et al. Genetic diversity and population structure of the upriver orange mangrove Bruguiera sexangula along the coastlines of Thailand[J]. Aquat Biol,2023,32:31−41. doi: 10.3354/ab00761
[22] Zhang Y,Xin K,Sheng N,Xie ZL,Liao BW. The regenerative capacity of eight mangrove species based on propagule traits in Dongzhai Harbor,Hainan Province,China[J]. Glob Ecol Conserv,2021,30:e01788.
[23] Xin K,Xie ZL,Zhong CR,Sheng N,Gao C,et al. Damage caused by Sphaeroma to Mangrove Forests in Hainan,Dongzhaigang,China[J]. J Coastal Res,2020,36(6):1197−1203.
[24] Qayoom H,Alkhanani M,Almilaibary A,Alsagaby SA,Mir MA. A network pharmacology-based investigation of Brugine reveals its multi-target molecular mechanism against breast cancer[J]. Med Oncol,2023,40(7):202. doi: 10.1007/s12032-023-02067-w
[25] 林鹏,卢昌义,王恭礼,陈焕雄. 海南岛河港海莲红树林凋落物动态的研究[J]. 植物生态学与地植物学学报,1990,14(1):69−74. Lin P,Lu CY,Wang GL,Chen HX. Study on dynamics of litter fall of Bruguiera sexangula mangrove in Hainan Island,China[J]. Acta Phytoecologyet Geobotanica Sinica,1990,14(1):69−74.
[26] Li CM,Wang HC,Liao XL,Xiao R,Liu KH,et al. Heavy metal pollution in coastal wetlands:a systematic review of studies globally over the past three decades[J]. J Hazard Mater,2022,424:127312. doi: 10.1016/j.jhazmat.2021.127312
[27] Niu YH,An ZR,Gao DZ,Chen FY,Zhou J,et al. Tidal dynamics regulates potential coupling of carbon-nitrogen-sulfur cycling microbes in intertidal flats[J]. Sci Total Environ,2023,899:165663. doi: 10.1016/j.scitotenv.2023.165663
[28] Yu ZW,Wang ML,Sun ZY,Wang WJ,Chen Q. Changes in the leaf functional traits of mangrove plant assemblages along an intertidal gradient in typical mangrove wetlands in Hainan,China[J]. Glob Ecol Conserv,2023,48:e02749.
[29] 吕晓波,钟才荣,张孟文,方赞山,程成,等. 海南东寨港红海榄种群结构与数量动态变化特征[J]. 热带生物学报,2023,14(5):490−498. Lü XB,Zhong CR,Zhang MW,Fang ZS,Cheng C,et al. Population structure and dynamics of Rhizophora stylosa in Dongzhaigang National Mangrove Reserve,Hainan[J]. Journal of Tropical Biology,2023,14(5):490−498.
[30] 辜晓虹,邱彭华,陈卫,周文芊,陈晓娟,杨仕莉. 海口东寨港自然保护区红树林变化及其与周边社区关联分析[J]. 热带地理,2023,43(1):43−58. Gu XH,Qiu PH,Chen W,Zhou WQ,Chen XJ,Yang SL. Mangrove forest change in Haikou Dongzhaigang Nature Reserve and its association with surrounding communities[J]. Tropical Geography,2023,43(1):43−58.
[31] 张育霞. 不同潮间带角果木种群特征及其繁殖特性研究[D]. 海口:海南大学,2015:11−14. [32] 何东进,肖石红,李晓景,王韧,蔡金标,游巍斌. 中国红树林北缘区秋茄树种群空间分布格局和结构特征分析[J]. 植物资源与环境学报,2014,23(2):41−47. doi: 10.3969/j.issn.1674-7895.2014.02.06 He DJ,Xiao SH,Li XJ,Wang R,Cai JB,You WB. Analyses on spatial distribution pattern and structure characteristics of Kandelia candel population in north marginal region of mangrove,China[J]. Journal of Plant Resources and Environment,2014,23(2):41−47. doi: 10.3969/j.issn.1674-7895.2014.02.06
[33] 江洪. 云杉种群生态学[M]. 北京:中国林业出版社,1992:7−13. [34] 王进,姚兰,艾训儒,朱江,刘松柏. 鄂西南不同区域亮叶桦种群结构与动态特征[J]. 应用生态学报,2020,31(2):357−365. Wang J,Yao L,Ai XR,Zhu J,Liu SB. Structure and dynamic characteristics of Betula luminifera populations in different regions of Southwest Hubei Province,China[J]. Chinese Journal of Applied Ecology,2020,31(2):357−365.
[35] 牛翠娟,娄安如,孙儒泳,李庆芬. 基础生态学[M]. 北京:高等教育出版社,2015:151−152. [36] Shaltout KH,Ahmed DA,Shabana HA. Population structure and dynamics of the endemic species Phlomis aurea Decne in different habitats in southern Sinai Peninsula,Egypt[J]. Glob Ecol Conserv,2015,4:505−515.
[37] 李大东,董廷发,陈坚,史清茂,何显湘,等. 四川米仓山自然保护区台湾水青冈群落学特征及多样性研究[J]. 西北植物学报,2016,36(1):174−182. Li DD,Dong TF,Chen J,Shi QM,He XX,et al. Characteristics of Fagus hayatae community and species diversity in Micangshan Nature Reserve,Sichuan[J]. Acta Botanica Boreali-Occidentalia Sinica,2016,36(1):174−182.
[38] Lefcheck JS,Byrnes JEK,Isbell F,Gamfeldt L,Griffin JN,et al. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats[J]. Nat Commun,2015,6(1):6936. doi: 10.1038/ncomms7936
[39] 廖宝文. 三种红树植物对潮水淹浸与水体盐度适应能力的研究[D]. 北京:中国林业科学研究院,2010:111−118. [40] 陈鹭真,林鹏,王文卿. 红树植物淹水胁迫响应研究进展[J]. 生态学报,2006,26(2):586−593. Chen LZ,Lin P,Wang WQ. Mechanisms of mangroves waterlogging resistance[J]. Acta Ecologica Sinica,2006,26(2):586−593.
[41] Deborde J,Marchand C,Molnar N,Della Patrona L,Meziane T. Concentrations and fractionation of carbon,iron,sulfur,nitrogen and phosphorus in mangrove sediments along an intertidal gradient (semi-arid climate,New Caledonia)[J]. J Mar Sci Eng,2015,3(1):52−72. doi: 10.3390/jmse3010052
[42] 林鹏. 中国东南部海岸红树林的类群及其分布[J]. 生态学报,1981,1(3):283−290. Lin P. Ecological notes on mangroves in Southeast coast of China including Taiwan Province and Hainan Island[J]. Acta Ecologica Sinica,1981,1(3):283−290.
[43] 陈玉军,李婷,朱立安,林梓,贾桐,朱虎. 淹水梯度对红树林湿地土壤微生物生物量与酶活性的影响[J]. 水土保持通报,2022,42(6):68−75. Chen YJ,Li T,Zhu LA,Lin Z,Jia T,Zhu H. Effects of flooding gradient on soil microbial biomass and enzyme activities in a mangrove wetland[J]. Bulletin of Soil and Water Conservation,2022,42(6):68−75.
[44] 詹有生. 江西部分常绿阔叶林物种多样性的研究[J]. 南京林业大学学报,1997,21(1):67−70. Zhan YS. Study on the species diversity in part of the evergreen broad leaved forest communities of Jiangxi[J]. Journal of Nanjing Forestry University,1997,21(1):67−70.
[45] 彭宗波,蒋英. 优势种密度制约及其对森林群落物种多样性维持的影响[J]. 资源与生态学报,2016,7(4):275−280. doi: 10.5814/j.issn.1674-764x.2016.04.006 Peng ZB,Jiang Y. Density dependence of a dominant species and the effects on community diversity maintainance[J]. Journal of Resources and Ecology,2016,7(4):275−280. doi: 10.5814/j.issn.1674-764x.2016.04.006
[46] Hacker SD,Bertness MD. Experimental evidence for factors maintaining plant species diversity in a New England salt marsh[J]. Ecology,1999,80(6):2064−2073. doi: 10.1890/0012-9658(1999)080[2064:EEFFMP]2.0.CO;2
[47] 李大东,胥晓,史清茂,陈坚,昝玺,等. 四川米仓山自然保护区台湾水青冈群落学特征沿海拔梯度的变化[J]. 热带亚热带植物学报,2016,24(6):626−634. Li DD,Xu X,Shi QM,Chen J,Zan X,et al. Characteristics of Fagus hayatae community along altitudinal gradient in Micangshan Nature Reserve,Sichuan[J]. Journal of Tropical and Subtropical Botany,2016,24(6):626−634.
[48] McKee KL. Seedling recruitment patterns in a Belizean mangrove forest:effects of establishment ability and physico-chemical factors[J]. Oecologia,1995,101(4):448−460. doi: 10.1007/BF00329423
[49] 吕晓波. 海南岛六种红树植物幼苗对光照、盐度和淹水时间的生理适应性研究[D]. 海口:海南大学,2019:218−227. [50] 王立龙,王亮,张丽芳,刘玉洋,徐世健. 不同生境下濒危植物裸果木种群结构及动态特征[J]. 植物生态学报,2015,39(10):980−989. doi: 10.17521/cjpe.2015.0095 Wang LL,Wang L,Zhang LF,Liu YY,Xu SJ. Structure and dynamic characteristics of Gymnocarpos przewalskii in different habitats[J]. Chinese Journal of Plant Ecology,2015,39(10):980−989. doi: 10.17521/cjpe.2015.0095
[51] 李敏敏,刘鹏程,孔维民,马方莲,李帅锋,王萌. 濒危植物澜沧黄杉种群结构及动态特征[J]. 生态学报,2022,42(13):5504−5515. Li MM,Liu PC,Kong WM,Ma FL,Li SF,Wang M. Population structure and dynamic characteristics of the endangered Pseudotsuga forrestii Craib[J]. Acta Ecologica Sinica,2022,42(13):5504−5515.
[52] 刘震,闫淑君,田高飞,吴艳芳,段嵩岚,潘刚. 小果紫薇林近自然种群结构及动态特征[J]. 森林与环境学报,2017,37(3):342−347. Liu Z,Yan SJ,Tian GF,Wu YF,Duan SL,Pan G. Structure and dynamic characteristics of Lagerstroemia minuticarpa near-natural population[J]. Journal of Forest and Environment,2017,37(3):342−347.
计量
- 文章访问数: 283
- HTML全文浏览量: 90
- PDF下载量: 20
