
Karyotype of five Astragalus species from the alpine subnival belt in the Hengduan Mountains
-
摘要:
通过常规植物染色压片法对采自横断山高山冰缘带地区豆科黄耆属(Astragalus)5种植物进行了染色体数目及核型分析。结果显示:川西黄耆(A. craibianus Simps.)、无茎黄耆(A. acaulis Baker)、梭果黄耆(A. ernestii Comb.)、东俄洛黄耆(A. tongolensis Ulbr.)为二倍体,窄翼黄耆(A. degensis Ulbr.)为四倍体,所有物种的染色体基数均为x = 8;川西黄耆的染色体核型公式为2n = 2x = 16 = 6sm + 10m,为2A型,无茎黄耆的染色体核型公式为2n = 2x = 16 = 2sm + 14m,为2A型,梭果黄耆的染色体核型公式为2n = 2x = 16 = 2sm + 14m,为2A型,窄翼黄耆的染色体核型公式为2n = 4x = 32 = 4sm + 28m,为1A型,东俄洛黄耆的染色体核型公式为2n = 2x = 16 = 2st + 2sm + 12m,为2A型。已报道横断山的黄耆属物种均为二倍体,而本研究首次发现了四倍体种,研究结果补充了横断山高山地区黄耆属植物染色体的基础资料,同时为该区域的植物系统分类及属内植物进化研究提供了细胞分类学上的参考。
Abstract:The chromosome numbers and karyotypes of five species of Astragalus (Fabaceae) collected from the alpine subnival belt in the Hengduan Mountains were determined. All species were analyzed by conventional staining and pressing methods. The karyotype formulae of the five species are reported for the first time. Results showed that: A. craibianus Simps., A. acaulis Baker, A. ernestii Comb., A. tongolensis Ulbr. were diploid, while A. degensis Ulbr. was tetraploid. The chromosome base number for all studied species was x = 8. The karyotype formula of A. craibianus was 2n = 2x = 16 = 6sm + 10m, 2A. The karyotype formula of A. acaulis was 2n = 2x = 16 = 2sm + 14m, 2A. The karyotype formula of A. ernestii was 2n = 2x = 16 = 2sm + 14m, 2A. The karyotype formula of A. degensis was 2n = 4x = 32 = 4sm + 28m, 1A. The karyotype formula of A. tongolensis was 2n = 2x = 16 = 2st + 2sm + 12m, 2A. All species of Astragalus in the Hengduan Mountains were previously reported to be diploid, but this study discovered a tetraploid species for the first time. These results not only expand basic information on the chromosomes of legumes in the alpine subnival belt of the Hengduan Mountains, but also provide a cytological reference for the study of plant phylogeny and evolution of plants within Astragalus in this region.
-
Keywords:
- Hengduan Mountains /
- Astragalus /
- Chromosome number /
- Karyotype analysis
-
种间联结指的是不同物种间在特定空间分布中的相互关系[1],它是植物群落结构形成和变化的重要基础[2]。通过对群落种间联结的研究,能够确定植物的种间关系和预测种群消长动态,揭示群落中植物替代关系机制[3]。群落稳定性指的是群落外部条件改变或存在不同程度的扰动时,群落生态系统保持恒定状态的能力[4],它是植物群落结构与功能的一个综合特征,研究群落稳定性有利于认识森林群落的维持机制和演替规律[5]。对群落种间联结及群落稳定性的研究不仅能清晰地认识群落结构和功能,而且可为濒危种群恢复和生物多样性保护等工作的展开提供理论依据[6]。
金花茶组(Sect. Chrysantha)为山茶科山茶属(Camellia)常绿灌木,花为金黄色,是国家珍稀濒危植物[7]。针对金花茶组植物开展的研究包括:种群分布[8]、生理特征[9]、种质资源[10]、就地保护[11]等方面,为该组植物资源的保护和利用提供了有益的科学依据。但由于金花茶组自身存有结实率低、种子与花粉难传播等问题,致使其繁殖更新困难,也是金花茶组植物濒危的重要原因[12],再加上近年来人们因经济利益对野生金花茶非法采挖,已严重威胁野外金花茶组植物种群的发展[13],所以现阶段金花茶组植物的保护仍有不少问题,也反映出对其保护的必要性与重要性。2021年,金花茶组全部物种均被列入《国家重点保护野生植物名录》。
淡黄金花茶(C. flavida Hung T. Chang)隶属于金花茶组植物,花呈淡黄色,主要分布于广西崇左[14],其属耐阴灌木,具有较高的观赏和科研价值[15]。研究表明,淡黄金花茶对生境有喜好性且分布较为狭窄、零散,导致原生种数量锐减[16],这给淡黄金花茶的保护工作带来了困扰。目前,众学者多以研究淡黄金花茶的生物学特性为主,但本质仍为种内关系的研究,易忽视与植物群落内的关联,且淡黄金花茶与群落各物种有着不同的关系,也受群落动态影响,故对其种间关系及群落稳定性进行探究,将利于揭示该种群生态习性与植物群落的内在联系。在弄岗保护区中保存有较为完整的淡黄金花茶自然群落[16],因此,本研究以2011和2021年(间隔10年)两次淡黄金花茶伴生群落调查数据为基础,基于方差比率法(VR)、
χ2 检验、Spearman秩相关检验和M.Godron稳定性测定等方法,对群落中乔木层、灌木层优势种的种间联结及群落稳定性进行分析,拟解决以下科学问题:(1)淡黄金花茶伴生群落不同层次优势种的种间关系如何?(2)伴生群落特征及其稳定性对淡黄金花茶种群的保育有何影响?研究结果旨在探明淡黄金花茶伴生群落优势物种在群落中的地位,群落的发展动态及演替的趋势,并为制定淡黄金花茶的保护策略及深入研究提供理论基础。1. 材料与方法
1.1 研究区概况
广西弄岗国家级自然保护区位于龙州、宁明两县交界处(22.23°~22.55°N, 106.71°~107.08°E),呈东南至西北长条状分布,总面积约10 080 hm2。该区属热带季风气候,年均气温约22 ℃,其中,最冷月均温13 ℃,最热月均温28 ℃;年均降水量为1150~1550 mm,大多集中于6-8月,干湿交替季节明显,全年无霜期约351 d;土壤以黑色和棕色石灰土为主[17]。
弄岗保护区植物资源丰富,目前共记录蕨类植物有29科51属151种,裸子植物4科5属11种,被子植物142 科727 属1551种。国家一级保护植物有望天树(Parashorea chinensis H. Wang)、海伦兜兰(Paphiopedilum helenae Aver)等 6种;国家二级保护植物有蚬木(Excentrodendron tonkinense (A. Chev.) H. T. Chang & R. H. Miao)、东京桐(Deutzianthus tonkinensis Gagnep)、大叶风吹楠(Horsfieldia kingi (Hook. f.) Warb)等49种[18]。
保护区仍保存着原生性热带喀斯特季节性雨林,区内地貌主要为典型的喀斯特峰丛洼地,从洼地到山顶仅200~300 m的高度差,但生境异质性极强,发育形成了3种类型的生境。洼地周围分布着喜湿耐荫性的树种,以望天树、大叶风吹楠、中国无忧花(Saraca dives Pierre)等为优势树种;中坡周围分布着具有旱生特征的树种,以喀斯特专性植物蚬木、肥牛树(Cephalomappa sinensis (Chun & F. C. How) Kosterm)、金丝李(Garcinia paucinervis Chun ex F. C. How)等为优势树种;山顶周围分布着强耐旱性的树种,以清香木(Pistacia weinmanniifolia J. Poiss. ex Franch)、细叶谷木(Memecylon scutellatum (Lour.) Hook. & Arn)、黄梨木(Boniodendron minus (Hemsl.) T. Chen)等为优势树种。
1.2 研究方法
1.2.1 样地设置与群落调查
2011年,在保护区中淡黄金花茶发育完好且分布集中的洼地区域设置10个20 m×30 m的样方,每个样方再分为6个10 m×10 m的小样格,进行乔木层和灌木层的植物调查,乔木层、灌木层的划分标准参考中国植被志群落调查规范[19]。调查时,对乔木层样方内所有胸径大于3 cm的乔木进行每木调查,测量并记录种名、坐标、胸径和生长状态等信息;对灌木层内所有灌木个体(包含乔木幼树)记录种名、坐标、基径和生长状态等信息。于2021年开展植物群落复查,详细记录乔木层、灌木层植物胸径、基径、存活状态等信息。
1.2.2 重要值计算
选择两次调查中乔木层和灌木层中重要值都大于2%的优势种展开种间联结分析。同时,将淡黄金花茶加入乔木层进行分析,故乔木层12种,灌木层15种。重要值公式[4]如下:
IV(乔木)=(相对密度+相对频度+相对优势度)/3 IV(灌木)=(相对密度+相对频度)/2 相对密度(%)=(某个种的株数/所有种的总株数)×100 相对频度(%)=(某个种的频度/所有种的频度之和)×100 相对优势度(%)=(某个种的胸高断面积之和/所有种的胸高断面积之和)×100 1.2.3 总体联结性检验
总体联结性采用方差比率(VR)[20]来测定,并通过统计量(W)检验总体联结性是否显著,公式[21]如下:
VR=S2T/δ2T=1NN∑j=1(Tj−t)2/S∑i=1niN(1−niN) 式中,
S2T 为总种数方差,δ2T 为总体样本方差,S为总物种数,N为样方总数,ni为物种i出现的样方数,Tj为样方j中出现的物种数,t为所有样方中物种数的平均值。VR表示总体联结性指数,若VR=1,表示无联结;VR>1,表示正联结;VR<1,表示负联结。同时,需通过统计量W(W=N× VR)检验VR值是否显著,若统计量(W)落入效应区间:χ20.95 (N)<W<χ20.05 (N),表示联结不显著,反之则表示联结显著。1.2.4 种间联结性
种间联结性的
χ2 统计量通过Yates连续校正公式计算,公式[21]如下:χ2=N(|ad−bc|−0.5N)2(a+b)(a+c)(b+d)(c+d) 式中,N为样方总数,a为两物种的同时出现样方数,b和c为任一物种单独出现的样方数,d为两物种均未出现的样方数。
χ2 >6.635,表示种间联结极显著;若3.841≤χ2 ≤6.635,则表示种间联结显著;若χ2 <3.841,则表示种间联结不显著。用V值判断种间正负联结性,V=[(a+b)−(b+c)]/(a+b+c+d),当V>0时为正联结;当V<0时为负联结。1.2.5 种间相关性
采取Spearman秩相关系数来判断不同层次优势种对间的相关性。公式[4]如下:
rs(i,j)=1−[6N∑k=1(xik−xjk)2/(N3−N)] 式中, rs(i,j)是物种i和物种j在样方k中的Spearman秩相关系数,值域为[−1,1],正值为正相关,负值为负相关,N为样方总数,xik和xjk表示物种i和物种j在样方k中的秩。
1.2.6 群落稳定性
本研究采用郑元润改进后的M. Godron稳定性测定法[22],以群落物种累计倒数百分比和累积相对频度分别作为x轴与y轴,用方程( y=ax2+bx+c)拟合,制作平滑曲线连接的散点图,使其与直线 y=100−x相交,交点坐标即群落稳定性比值,其越接近(20,80),说明群落越稳定,反之则越不稳定。
1.3 数据处理
本研究采用Excel 2021软件计算群落重要值和群落稳定性,而方差比率 (VR)、
χ2 检验、Spearman秩相关检验等在R4.2.2软件的spaa包[23]中完成。2. 结果与分析
2.1 群落特征与优势物种重要值变化
淡黄金花茶伴生群落分布于喀斯特季节性雨林的洼地周围,洼地伴有季节性水淹,土壤湿度大,群落木本植物垂直结构成层性明显,可分为乔木层与灌木层,乔木层以中国无忧花、广西棋子豆(Archidendron guangxiensis T. L. Wu)等优势种占据群落上层,调查到群落植株的最高高度达32 m,乔木层10年间植株平均胸径从7.99 cm增至9.05 cm,且乔木层多数乔木冠幅较大,遮蔽了群落上层绝大部分阳光,为下层灌木及乔木幼树创造了遮荫的生境,而灌木层以对叶榕(Ficus hispida L. f.)、南方紫金牛(Ardisia thyrsiflora D. Don) 等优势种占据乔木层林下,灌木层10年间植株平均胸径从1.67 cm增至1.85 cm,植株平均高度为2~3 m,与乔木层有明显的分层现象。群落种类组成丰富,据统计,2011和2021年分别调查到植物种类为45科86属103种和41科70属86种,种类较多的科有叶下珠科、桑科和楝科等。
伴生群落乔木层、灌木层优势种重要值的变化见表1,2011-2021年间,乔木层与灌木层优势种在群落中的重要值变化幅度相对较小(1%左右),说明这些优势种在群落中生存状态稳定。其中淡黄金花茶在不同层次的重要值均有提升,说明随着时间的进行该生境越利于淡黄金花茶种群的生存与发展。
表 1 淡黄金花茶伴生群落不同年份乔木层、灌木层优势种及其重要值Table 1. Dominant species and their importance values in tree and shrub layers of Camellia flavida-associated community across different years优势物种
Dominant species缩写
Abbreviation重要值
Importance value / %2011 2021 乔木层 苹婆Sterculia monosperma Vent Sm 17.89 17.17 中国无忧花Saraca dives Pierre Sd 10.73 11.58 广西牡荆Vitex kwangsiensis C. Pei Vk 7.85 7.55 广西棋子豆Archidendron guangxiensis T. L. Wu Ag 7.22 7.75 对叶榕Ficus hispida L. f. Fh 4.67 6.68 假肥牛树Cleistanthus petelotii Merr. ex Croizat Cp 4.63 5.32 劲直刺桐Erythrina stricta Roxb Es 3.63 2.93 苹果榕Ficus oligodon Miq Fo 3.32 3.21 肥牛树Cephalomappa sinensis (Chun & F. C. How) Kosterm Cs 3.19 2.59 日本五月茶Antidesma japonicum Siebold & Zucc Aj 3.14 3.46 海南大风子Hydnocarpus hainanensis (Merr.) Sleumer Hh 2.82 3.27 淡黄金花茶 Camellia flavida Hung T. Chang Cf 0.31 1.32 灌木层 对叶榕Ficus hispida L. f. Fh 10.28 8.21 日本五月茶Antidesma japonicum Siebold & Zucc Aj 9.35 11.08 假肥牛树Cleistanthus petelotii Merr. ex Croizat Cp 6.66 7.14 南方紫金牛Ardisia thyrsiflora D. Don At 5.78 6.98 苹婆Sterculia monosperma Vent Sm 5.23 3.71 山桂花Bennettiodendron leprosipes (Clos) Merr Bl 5.14 6.99 淡黄金花茶 Camellia flavida Hung T. Chang Cf 4.35 6.17 广西棋子豆 Archidendron guangxiensis T. L. Wu Ag 3.85 5.08 广西澄广花Orophea polycarpa A. Candolle Op 3.83 3.27 垂茉莉Clerodendrum wallichii Merr Cw 3.70 4.21 海南大风子Hydnocarpus hainanensis (Merr.) Sleumer Hh 3.57 4.72 金丝李Garcinia paucinervis Chun ex F. C. How Gp 2.55 2.89 茎花山柚Champereia manillana var. longistaminea (W. Z. Li) H. S. Kiu Cm 2.31 2.14 南烛厚壳桂Cryptocarya lyoniifolia S. Lee et F. N. Wei Cl 2.30 2.19 裂果卫矛Euonymus dielsianus Loes. & Diels Ed 2.15 2.67 2.2 总体联结性变化
淡黄金花茶伴生群落中乔木层、灌木层优势种间的总体联结性特征见表2,从2011-2021年乔木层优势种方差比率(VR)均小于1,且检验统计量(W)均落入效应区间,表明乔木层优势种的总体联结性呈不显著负联结;灌木层优势种方差比率(VR)均大于1,且检验统计量(W)均未落入效应区间,表明灌木层优势种的总体联结性呈显著正联结。乔木层和灌木层的方差比率值均有所增加,说明随时间推移,乔木层逐渐趋于正联结,灌木层正联结程度不断增强。
表 2 淡黄金花茶群落伴生群落乔木层和灌木层优势种不同年份总体联结性检验Table 2. Overall interspecific associations among dominant tree and shrub layer species in Camellia flavida-associated community across different years层次
Level年份
Year方差比率
Variance ratio (VR)统计量(W)
Statistic (W)χ2临界检验值
χ2 threshold结果
Results乔木层 2011 0.71 43.49 (43.19,79.08) 不显著负联结 2021 0.90 54.22 (43.19,79.08) 不显著负联结 灌木层 2011 1.54 92.49 (43.19,79.08) 显著正联结 2021 1.60 95.61 (43.19,79.08) 显著正联结 2.3 种间联结性变化
2.3.1 乔木层种间联结性变化
2011和2021年乔木层优势种
χ2 检验比较结果显示:在优势种组成的66个种对中,正联结种对由36对变为32对,占总种对数比由54.55%变为48.48%;负联结种对不变,为26对,占总种对数比为39.39%;无联结种对由4对变为8对,占总种对数比由6.06%变为12.12%。其中,淡黄金花茶与海南大风子(Hydnocarpus hainanensis (Merr.) Sleumer)呈极显著负联结,与对叶榕、中国无忧花呈显著负联结,而与苹果榕呈极显著正联结。从总体来看,随时间变化乔木层负联结种对变化不明显,正联结种对减少,无联结种对增加(图1)。![]() 图 1 淡黄金花茶伴生群落不同年份乔木层优势种
图 1 淡黄金花茶伴生群落不同年份乔木层优势种χ2 检验半矩阵图★:极显著正联结;▲:显著正联结;+:不显著正联结;☆:极显著负联结;△:显著负联结;-:不显著负联结;○:无联结。下同。Figure 1. Semi-matrix diagram ofχ2 test results of interspecific associations among dominant tree layer species in Camellia flavida-associated community across different years★: Extremely significant positive association; ▲: Significant positive association; +: No significant positive association; ☆: Extremely significant negative association; △: Significant negative association; -: No significant negative association; ○: No association. Same below.2.3.2 灌木层种间联结性变化
2011和2021年灌木层优势种
χ2 检验比较结果显示(图2):在优势种组成的105个种对中,正联结种对由66对变为77对,占总种对数比由62.86%变为73.33%;负联结种对由37对变为25对,占总种对数比由35.24%变为23.81%;不显著联结和无联结种对无变化,均为93对,占总种对数比88.57%。其中,淡黄金花茶与灌木层大多数物种呈正联结。综合χ2 检验结果,发现灌木层正联结种对随着时间变化明显增多,负联结种对数量逐渐减少,这与总体联结性检验结果相似,说明随群落的演替灌木层正联结程度逐渐增强。![]() 图 2 淡黄金花茶伴生群落不同年份灌木层优势种
图 2 淡黄金花茶伴生群落不同年份灌木层优势种χ2 检验半矩阵图Figure 2. Semi-matrix diagram ofχ2 test results for interspecific associations among dominant shrub layer species in Camellia flavida-associated community across different years2.4 种间相关性变化
2.4.1 乔木层种间相关性变化
乔木层优势物种Spearman秩相关检验结果表明,从2011-2021年,在优势种组成的66个种对中,正相关种对由27对变为30对,占总种对数比由40.91%变为45.45%,其中负相关种对由39对变为36对,占总种对数比由59.09%变为54.55%,其中显著相关种对由27对变为28对,占总种对数比由40.91%变为42.42%。该层次中,淡黄金花茶相关性检验结果与
χ2 检验相似,其与对叶榕、海南大风子及中国无忧花等表现为显著负相关,而与日本五月茶(Antidesma japonicum Siebold & Zucc.)、苹果榕和假肥牛树(Cleistanthus petelotii Merr. ex Croizat)呈显著正相关。总的看来,Spearman秩相关检验显著率大于χ2 检验,且随着时间的推移,正相关种对逐渐增加,负相关种对逐渐减少(图3)。![]() 图 3 淡黄金花茶伴生群落不同年份乔木层优势种Spearman秩相关检验半矩阵图Figure 3. Semi-matrix diagram of Spearman rank correlation coefficients among dominant tree layer species in Camellia flavida-associated community across different years*: P<0.05; **: P<0.01; ***: P<0.001. Same below.
图 3 淡黄金花茶伴生群落不同年份乔木层优势种Spearman秩相关检验半矩阵图Figure 3. Semi-matrix diagram of Spearman rank correlation coefficients among dominant tree layer species in Camellia flavida-associated community across different years*: P<0.05; **: P<0.01; ***: P<0.001. Same below.2.4.2 灌木层种间相关性变化
灌木层优势种Spearman秩相关检验结果表明(图4),从2011-2021年,在优势种组成的105个种对中,正相关种对由65对变为59对,占总种对数比由61.90%变为56.19%;负相关种对由40对变为46对,占总种对数比由38.10%变为43.81%;其中显著相关种对由25对变为29对,占总种对数比由23.81%变为27.62%。其中,十年间淡黄金花茶仅与垂茉莉(Clerodendrum wallichii Merr)和日本五月茶呈显著负相关,与广西澄广花、南烛厚壳桂(Cryptocarya lyoniifolia S. Lee et F. N. Wei)呈显著正相关,而与其他物种之间大多相关性不显著。
![]() 图 4 淡黄金花茶伴生群落不同年份灌木层优势种Spearman秩相关检验半矩阵图Figure 4. Semi-matrix diagram of Spearman rank correlation coefficients among dominant shrub layer species in Camellia flavida-associated community across different years
图 4 淡黄金花茶伴生群落不同年份灌木层优势种Spearman秩相关检验半矩阵图Figure 4. Semi-matrix diagram of Spearman rank correlation coefficients among dominant shrub layer species in Camellia flavida-associated community across different years2.5 群落稳定性变化
2011年与2021年群落稳定性分析比较结果显示(表3、图5),乔木层、灌木层及群落整体的稳定性比值均相对靠近于(20,80),且各层次及整体的交点坐标变化不大,说明淡黄金花茶伴生群落较为稳定。
表 3 淡黄金花茶伴生群落不同年份稳定性分析结果Table 3. Camellia flavida-associated community stability across different years年份
Year层次
Level曲线类型
Type of curve决定系数(R2)
Determination
coefficient交点坐标
Intersection coordinate2011 乔木层 y=−0.0131x2+1.977 7x+25.099 0.96 (28.80,71.20) 灌木层 y=−0.0143x2+2.122 1x+22.220 0.94 (28.68,71.32) 群落整体 y=−0.0142x2+2.110 8x+22.245 0.95 (28.77,71.23) 2021 乔木层 y=−0.0134x2+1.985 2x+27.118 0.95 (27.91,72.09) 灌木层 y=−0.0147x2+2.177 8x+20.107 0.95 (29.04,70.96) 群落整体 y=−0.0145x2+2.126 9x+23.487 0.94 (28.14,71.86) ![]() 图 5 淡黄金花茶伴生群落乔木层、灌木层不同年份稳定性Figure 5. Stability of tree and shrub layer in Camellia flavida-associated community across different years
图 5 淡黄金花茶伴生群落乔木层、灌木层不同年份稳定性Figure 5. Stability of tree and shrub layer in Camellia flavida-associated community across different years3. 讨论
植物群落中生态习性相近或生物特性相似的物种会表现出正联结关系,而生态特性差异或在竞争资源中相互排斥的物种会表现出负联结关系[24]。例如,本研究的淡黄金花茶与灌木层中的多数优势种呈正联结关系,原因是洼地分布的树种具喜湿耐荫性,且洼地区域更适幼树生长,个体竞争差异较小[25],这些相似的生物学特性与生态习性使得淡黄金花茶和灌木层其他种多表现为正联结;而在乔木层中淡黄金花茶与海南大风子、对叶榕及中国无忧花等树种呈不同程度的显著负联结,其中一原因有可能是海南大风子更适应较为干旱的中坡生境[26],与淡黄金花茶相互独立而呈极显著负联结,另一原因可能为对叶榕、中国无忧花等更适应潮湿的洼地生境,广泛分布于洼地周围且生态位宽度较大[26],故与其他物种间的生态位重叠程度也较高[1],导致了这些物种与淡黄金花茶表现出显著负联结关系。但淡黄金花茶存在种子重、花粉传播距离短等扩散限制问题,致使淡黄金花茶种群呈显著的聚集分布[27],使其和各层次优势物种的关系以不显著联结居多,这表明群落的种间联结性与物种的生物学特性、生态习性密切相关[28]。
种间联结理论认为,种间正相关是两物种在异质环境内因生存策略相似表现出相互依赖或相互兼容的关系,而负相关是由于两物种间对环境需求差异而发生相互排斥的关系[2]。例如,10年间淡黄金花茶与乔木层的苹果榕、灌木层的广西澄广花等优势种呈显著正相关,但其与乔木层的对叶榕、灌木层的日本五月茶等优势种呈显著负相关,原因可能为各优势种在喀斯特异质生境中所形成的种间关系不同,因为喀斯特季节性雨林洼地生境的土层较厚、土壤肥沃且潮湿[29],故不同生活型间种群聚集度较高[30],淡黄金花茶可能因相近的生存策略与苹果榕、广西澄广花等优势种互相依赖而表现出显著的正相关性,也可能因水、热资源竞争与对叶榕、日本五月茶等优势种相互排斥产生了显著的负相关。在实际调查中发现,淡黄金花茶喜欢生长于阴湿沟谷旁,这与乔木层、灌木层优势树种的生境偏好有一定差异,因此它与其他物种的相关性也稍弱。在喀斯特峰丛洼地中,异质生境促进了生态位高度分化[26],更多物种得以共存,群落组成变得复杂,令不同物种相遇概率变低,从而导致群落种间联结较为松散,相关性较弱。综上,部分优势乔灌木和喀斯特生境异质性很大程度上影响了淡黄金花茶的空间分布。
淡黄金花茶伴生群落位于保护核心区,良好的保护确保群落始终处于顶级群落阶段[27]。在伴生群落中,中国无忧花、广西棋子豆等高大树种占据乔木层,为林下创造了阴蔽的环境,且洼地的季节性水淹使得群落内较为潮湿,而这种阴蔽潮湿的生境更适宜淡黄金花茶种群的生长与繁育,10年的监测结果也证明这一点。从群落层次上看,10年间乔木层的平均胸径有所增长,说明乔木层处于动态更新状态,进而持续为淡黄金花茶提供遮荫的生存环境;反观灌木层10年间的平均胸径变化不大,说明灌木层相对稳定,在这种相同的层次与生境下,淡黄金花茶的重要值仍有提升,表明其在该生境下得到了良好的生存与发展。从群落种类组成上看,尽管伴生群落的丰富度减少,但减少种类多为群落数量稀少物种,而实际仍以优势乔灌木起主导作用,充分体现了伴生群落具有较高的稳定性,这为淡黄金花茶提供了稳定的群落环境,有利于其种群生存。由此看来,伴生群落特征及稳定性对淡黄金花茶种群的保育起到了积极的作用。
淡黄金花茶作为石灰岩专性植物,在喀斯特季节性雨林中分布相对狭窄、分散,但经过多年来弄岗保护区的严格保护,淡黄金花茶种群得到了良好的发展与恢复。通过10年的监测发现,伴生群落中胸径大于1 cm的淡黄金花茶个体数由68株增加至83株,植株数量呈增长趋势,说明保护区对于淡黄金花茶的就地保护工作取得较为显著的成效,故目前对其最好的保护与恢复策略就是继续加强就地保护,维持原生境的稳定性,避免人为干扰,通过自然恢复达到保护目的。
-
![]()
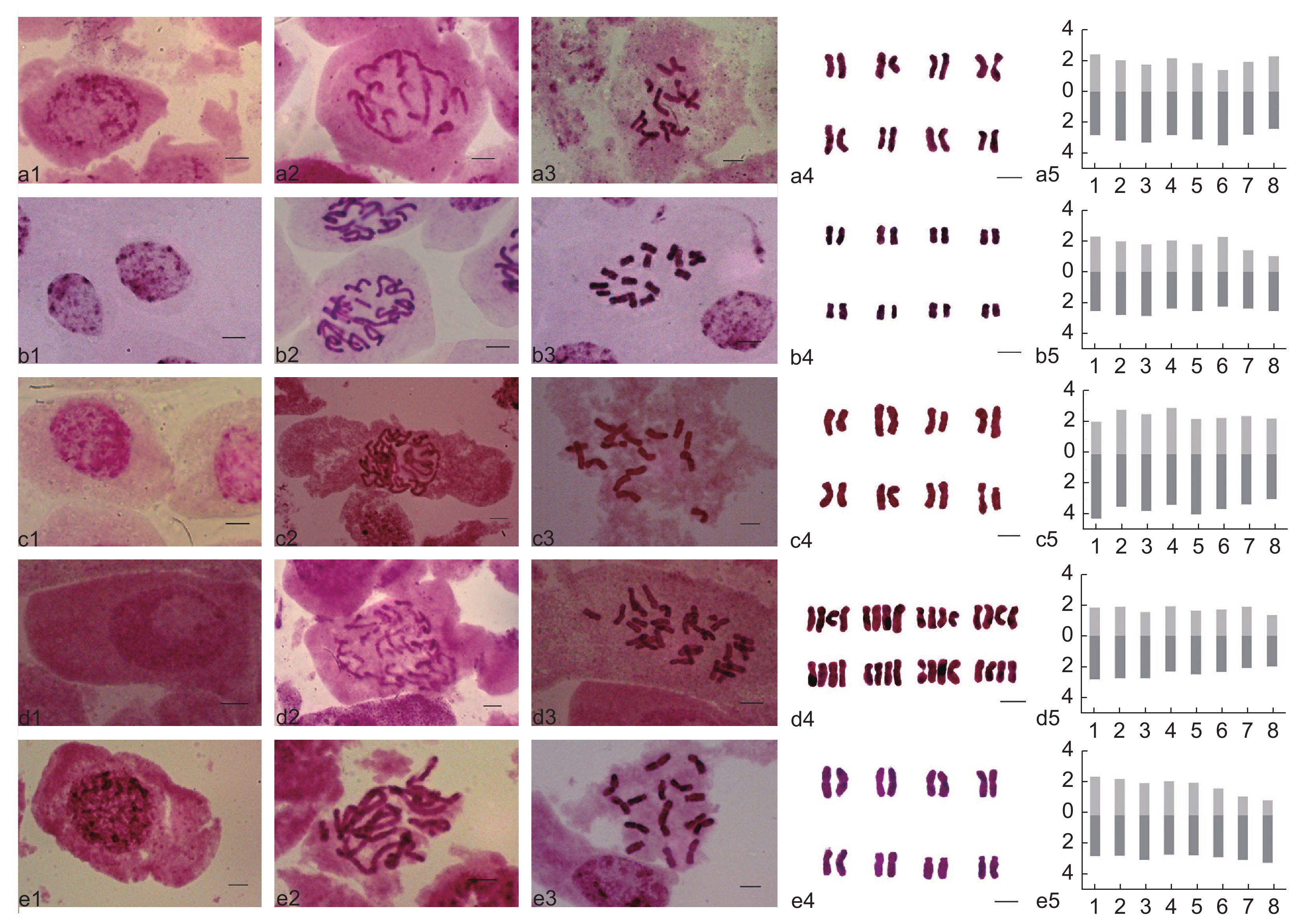
图 1 体细胞有丝分裂间期、前期、中期图、染色体配对图和核型模式图
a1~e1:体细胞有丝分裂间期核;a2~e2:体细胞有丝分裂前期图;a3~e3:体细胞有丝分裂中期染色体图;a4~e4:染色体抠图;a5~e5:核型模式图;a1~a5:川西黄耆;b1~b5:无茎黄耆;c1~c5:梭果黄耆;d1 ~d5:窄翼黄耆;e1~e5:东俄洛黄耆。(标尺 = 5 μm;a5~e5:x 轴为染色体的序号,y 轴为染色体的绝对长度;浅灰色为短臂,深灰色为长臂)。
Figure 1. Somatic mitotic interphase, prophase, metaphase chromosome, karyogram, and karyotype ideogram
a1-e1: Interphase; a2-e2: Prophase; a3-e3: Metaphase chromosomes; a4-e4: Karyogram; a5-e5: Karyotype ideogram; a1-a5: Astragalus craibianus; b1-b5: A. acaulis; c1-c5: A. ernestii; d1-d5: A. degensis; e1-e5: A. tongolensis. (Scale = 5 μm; a5-e5: x axis: Chromosome number, y-axis: Chromosome length; light gray: Short arm, dark gray: Long arm).
表 1 染色体已有核型研究
Table 1 Previous studies on chromosome karyotype
物种
Speciesx 2n 核型公式
Karyotype formula无茎黄耆
Astragalus acaulis Baker8 16 2n = 16[13] 梭果黄耆
Astragalus ernestii Comb.8 16 2n = 16[13] 东俄洛黄耆
Astragalus tongolensis Ulbr.8 16 2n = 16[13] 黑紫花黄耆
Astragalus przewalskii Bunge8 16 2n = 16[13] 白序黄耆
Astragalus leucocephalus R. Grah. ex Benth.8 16 2n = 16[14] 多花黄耆
Astragalus floridulus Podlech8
1616
322n = 16[13]
2n = 32[13]四川黄耆
Astragalus sutchuenensis Franch.8 16 2n = 2x = 16 = 8sm + 8m[10] 多枝黄耆
Astragalus polycladus Bur. et Franch.8 16 2n = 2x = 16 = 4sm + 12m(4SAT)[15] 密花黄耆
Astragalus densiflorus Kar. et Kir.8 16 2n =2x = 16 = 6m(2SAT) + 10sm[16]  下载: 导出CSV
下载: 导出CSV
表 2 实验所用材料采集地点信息
Table 2 Locations and vouchers of materials investigated
编号
No.物种
Species学名
Taxon采集地点
Location生境
Habitat海拔
Altitude / m经纬度
Longitude and latitude标本凭证号
Voucher specimens1 川西黄耆 Astragalus craibianus Simps. 云南省
德钦县流石滩 4292 28.42°N
99.00°EYNNU-19-296 2 无茎黄耆 Astragalus acaulis Baker 云南省
德钦县流石滩 4664 28.28°N
99.11°ESunWG-0291 3 梭果黄耆 Astragalus ernestii Comb. 四川省
乡城县流石滩 4309 29.14°N
100.28°EYNNU-19-177 4 窄翼黄耆 Astragalus degensis Ulbr. 云南省
德钦县流石滩 4065 28.42°N
99.00°EYNNU-19-367 5 东俄洛黄耆 Astragalus tongolensis Ulbr. 四川省
稻城县流石滩 3990 28.75°N
100.28°EYNNU-19-196
下载: 导出CSV
表 3 着丝点分类系统
Table 3 Centromere name system
命名
Name着丝点位置
Centromere site臂比值
RatioM 正中部着丝点 1.0 m 中部着丝点区 1.0 ~ 1.7 sm 亚中部着丝点区 1.7 ~ 3.0 st 亚端部着丝点区 3.0 ~ 7.0 t 端部着丝点区 7.0 ~ ∞ T 端部着丝点 ∞
下载: 导出CSV
表 4 染色体核型参数
Table 4 Karyotype and chromosomal index
物种
Speciesx 2n 平均着丝粒
指数
CI染色体平均
长度
CL不对称指数
AI核型不对称性
指数
As.K / %核型类型
KA核型公式
Karyotype formula川西黄耆 8 16 0.40 4.97 0.66 60.40 2A 2n = 2x = 16 = 6sm + 10m 无茎黄耆 8 16 0.41 4.32 1.58 58.80 2A 2n = 2x = 16 = 2sm + 14m 梭果黄耆 8 16 0.41 6.17 0.76 58.70 2A 2n = 2x = 16 = 2sm + 14m 窄翼黄耆 8 32 0.41 4.14 0.89 58.99 1A 2n = 4x = 32 = 4sm + 28m 东俄洛黄耆 8 16 0.41 4.84 1.96 58.75 2A 2n = 2x = 16 = 2st + 2sm + 12m
下载: 导出CSV
表 5 黄耆属5种植物染色体参数
Table 5 Chromosome parameters of five species of Astragalus
物种
Species编号
No.绝对长度 Absolute length 臂比
Cl染色体类型
Type长臂 L 短臂 S 总长 L + S 川西黄耆 1 2.43 2.28 4.71 1.07 m 2 2.84 2.41 5.25 1.18 m 3 2.84 2.14 4.98 1.33 m 4 2.80 1.94 4.74 1.44 m 5 3.20 2.02 5.22 1.59 m 6 3.13 1.83 4.96 1.70 sm 7 3.33 1.74 5.07 1.92 sm 8 3.50 1.38 4.88 2.58 sm 无茎黄耆 1 2.53 2.29 4.83 1.11 m 2 2.38 2.03 4.41 1.17 m 3 2.25 2.25 4.51 1.17 m 4 2.81 1.99 4.79 1.40 m 5 2.55 1.80 4.35 1.41 m 6 2.85 1.80 4.65 1.62 m 7 2.40 1.40 3.80 1.70 m 8 2.56 1.02 3.58 2.49 sm 梭果黄耆 1 3.37 3.06 6.43 1.10 m 2 3.52 2.93 6.44 1.19 m 3 2.30 2.37 5.37 1.26 m 4 3.34 2.51 5.85 1.32 m 5 3.78 2.64 6.43 1.42 m 6 3.67 2.40 6.07 1.52 m 7 3.96 2.33 6.29 1.67 m 8 4.33 2.15 6.47 2.04 sm 窄翼黄耆 1 2.08 1.86 3.94 1.12 m 2 2.31 1.91 4.22 1.21 m 3 2.35 1.69 4.03 1.39 m 4 2.74 1.86 4.59 1.47 m 5 1.98 1.33 3.32 1.49 m 6 2.51 1.6 4.11 1.56 m 7 2.82 1.79 4.61 1.58 m 8 2.74 1.53 4.27 1.79 sm 东俄洛黄耆 1 2.75 2.63 5.38 1.05 m 2 2.72 2.48 5.20 1.09 m 3 2.64 2.32 4.96 1.13 m 4 2.70 2.21 4.91 1.22 m 5 2.96 2.18 5.13 1.34 m 6 2.79 1.82 4.61 1.55 m 7 2.99 1.29 4.28 2.41 sm 8 3.20 1.04 4.24 3.19 st
下载: 导出CSV
-
[1] Wu ZY, Raven PH, Hong DY. Flora of China [DB/OL]. [2022-03-16]. www. iplant. cn/foc.
[2] 黎春花,刘国道. 豆科植物应用价值综述[J]. 热带农业科学,2008,28(4):75−80. Li CH,Liu GD. Multipurpose use of legumes[J]. Chinese Journal of Tropical Agriculture,2008,28 (4):75−80. Li CH, Liu GD. Multipurpose use of legumes[J]. Chinese Journal of Tropical Agriculture, 2008, 28(4): 75-80, 92.
[3] 中国科学院中国植物志编辑委员会. 中国植物志: 第42卷: 第1分册[M]. 北京: 科学出版社, 1993: 78. [4] Sanderson MJ,Wojciechowski MF. Diversification rates in atemperate legume clade:are there “so many species” of Astragalus (Fabaceae)[J]. Am J Bot,1996,83 (11):1488−1502. doi: 10.1002/j.1537-2197.1996.tb13942.x
[5] 中国科学院青藏高原综合科学考察队. 横断山区维管植物[M]. 北京: 科学出版社, 1993: 942-967. [6] Chen YS,Deng T,Zhou Z,Sun H. Is the East Asian flora ancient or not?[J]. Natl Sci Rev,2018,5 (6):920−932. doi: 10.1093/nsr/nwx156
[7] Myers N,Mittermeier RA,Mittermeier CG,Da Fonseca GAB,Kent J. Biodiversity hotspots for conservation priorities[J]. Nature,2000,403 (6772):853−858. doi: 10.1038/35002501
[8] 李懋学, 张敩方. 植物染色体研究技术[M]. 哈尔滨: 东北林业大学出版社, 1991: 1-20. [9] 王家坚,彭智邦,孙航,聂泽龙,孟盈. 青藏高原与横断山被子植物区系演化的细胞地理学特征[J]. 生物多样性,2017,25(2):218−225. Wang JJ,Peng ZB,Sun H,Nie ZL,Meng Y. Cytogeographic patterns of angiosperms flora of the Qinghai-Tibet plateau and Hengduan mountains[J]. Biodiversity Science,2017,25 (2):218−225. Wang JJ, Peng ZB, Sun H, Nie ZL, Meng Y. Cytogeographic patterns of angiosperms flora of the Qinghai-Tibet plateau and Hengduan mountains[J]. Biodiversity Science, 2017, 25(2): 218-225.
[10] 黎斌,常朝阳,吴振海,徐朗然. 国产6种黄耆属植物的核型研究[J]. 西北植物学报,2004,24(4):711−715. Li B,Chang ZY,Wu ZH,Xu LR. A karyotype study of six Astragalus species from China[J]. Acta Botanica Boreali-Occidentalia Sinica,2004,24 (4):711−715. Li B, Chang ZY, Wu ZH, Xu LR. A karyotype study of six Astragalus species from China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(4): 711-715.
[11] 王丽,顾志建,孙航. 青藏高原几种黄芪和棘豆植物核型的初步研究[J]. 云南植物研究,1994,16(1):53−59. Wang L,Gu ZJ,Sun H. Preliminary karyomorphological study on the plants in genera Oxytropis and Astragalus from Qinghai-Xizang Plateau[J]. Acta Botanica Yunnanica,1994,16 (1):53−59. Wang L, Gu ZJ, Sun H. Preliminary karyomorphological study on the plants in genera Oxytropis and Astragalus from Qinghai-Xizang Plateau[J]. Acta Botanica Yunnanica, 1994, 16(1): 53-59.
[12] 聂泽龙,孙航,顾志建. 横断山区被子植物染色体研究概况[J]. 云南植物研究,2004,26(1):35−57. Nie ZL,Sun H,Gu ZJ. A survey of chromosome numbers from Angiosperms of the Hengduan Mountains,S. W. China[J]. Acta Botanica Yunnanica,2004,26 (1):35−57. Nie ZL, Sun H, Gu ZJ. A survey of chromosome numbers from Angiosperms of the Hengduan Mountains, S. W. China[J]. Acta Botanica Yunnanica, 2004, 26(1): 35-57.
[13] Löve Á. Chromosome number reports LXXXIX[J]. Taxon,1985,34 (4):727−730. doi: 10.1002/j.1996-8175.1985.tb04449.x
[14] Ashraf M,Gohil RN. Studies on the cytology of Legumes of Kashmir Himalaya Ⅲ. Interpopulation differences in the karyotypes of 3 species of Astragalus L.[J]. Cytologia,1988,53 (3):543−549. doi: 10.1508/cytologia.53.543
[15] 刘玉红,王善敏. 多枝黄芪的核型研究[J]. 草地学报,1994,2(1):56−58. Liu YH,Wang SM. Studies on the karyotype of Astragalus polycladus[J]. Acta Agrestia Sinca,1994,2 (1):56−58. Liu YH, Wang SM. Studies on the karyotype of Astragalus polycladus[J]. Acta Agrestia Sinca, 1994, 2(1): 56-58.
[16] 黎斌,常朝阳,李思锋,徐朗然,陈彦生,郭晓思. 中国西北地区11种黄耆属植物的细胞学研究[J]. 西北植物学报,2002,22(3):467−475. Li B,Chang ZY,Li SF,Xu LR,Chen YS,Guo XS. Studies on cytology of 11 Astragalus species from Northwestern China[J]. Acta Botanica Boreali-Occidentalia Sinica,2002,22 (3):467−475. Li B, Chang ZY, Li SF, Xu LR, Chen YS, Guo XS. Studies on cytology of 11 Astragalus species from Northwestern China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2002, 22(3): 467-475.
[17] 徐波, 李志敏, 孙航. 横断山高山冰缘带种子植物[M]. 北京: 科学出版社, 2014: 153-155. [18] 刘亚辉,孟盈,杨永红,杨永平. 青藏高原六种棘豆属植物的染色体数目及核型报道[J]. 植物分类与资源学报,2011,33(4):423−431. Liu YH,Meng Y,Yang YH,Yang YP. Chromosome numbers and karyotypes of six Oxytropis species (Fabaceae) from the Qinghai-Tibetan Plateau,China[J]. Plant Diversity and Resources,2011,33 (4):423−431. Liu YH, Meng Y, Yang YH, Yang YP. Chromosome numbers and karyotypes of six Oxytropis species (Fabaceae) from the Qinghai-Tibetan Plateau, China[J]. Plant Diversity and Resources, 2011, 33(4): 423-431.
[19] 黎斌,李思锋,吴振海,常朝阳. 豆科山羊豆族2属5种植物的核型分析[J]. 西北植物学报,2007,27(9):1888−1891. Li B,Li SF,Wu ZH,Chang ZY. Karyotype of 5 leguminosae species in trib. Galegeae from northwest China[J]. Acta Botanica Boreali-Occidentalia Sinica,2007,27 (9):1888−1891. Li B, Li SF, Wu ZH, Chang ZY. Karyotype of 5 leguminosae species in trib. Galegeae from northwest China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2007, 27(9): 1888-1891.
[20] 黄荣福,沈颂东,卢学峰. 青藏高原东北部植物染色体数目和多倍性研究[J]. 西北植物学报,1996,16(3):310−318. Huang RF,Shen SD,Lu XF. Studies of the chromosome numbers and polyploidy for some plant in the North-East Qinghai-Xizang Plateau[J]. Acta Botanica Boreali-Occidentalia Sinica,1996,16 (3):310−318. Hung RF, Shen SD, Lu XF. Studies of the chromosome numbers and polyploidy for some plant in the North-East Qinghai-Xizang Plateau[J]. Acta Botanica Boreali-Occidentalia Sinica, 1996, 16(3): 310-318.
[21] 丁鸿,邱东萍,陈少雄. 植物染色体标本的制备和染色体核型分析研究进展[J]. 南方农业学报,2012,43(12):1958−1962. Ding H,Qiu DP,Chen SX. Research progress in plant chromosome samples preparation and karyotype analysis[J]. Journal of Southern Agriculture,2012,43 (12):1958−1962. Ding H, Qiu DP, Chen SX. Research progress in plant chromosome samples preparation and karyotype analysis[J]. Journal of Southern Agriculture, 2012, 43(12): 1958-1962.
[22] 李懋学,陈瑞阳. 关于植物核型分析的标准化问题[J]. 武汉植物学研究,1985,3(4):297−302. Li MX,Chen RY. A suggestion on the standardization of karyotype analysis in plants[J]. Journal of Wuhan Botanical Research,1985,3 (4):297−302. Li MX, Chen RY. A suggestion on the standardization of karyotype analysis in plants[J]. Journal of Wuhan Botanical Research, 1985, 3(4): 297-302.
[23] Altınordu F,Peruzzi L,Yu Y,He XJ. A tool for the analysis of chromosomes:KaryoType[J]. Taxon,2016,65 (3):586−592. doi: 10.12705/653.9
[24] Levan A,Fredga K,Sandberg AA. Nomenclature for centromeric position on chromosomes[J]. Hereditas,1964,52 (2):201−220.
[25] Stebbins GL. Chromosomal Evolution in Higher Plants[M]. London: Edward Arnold, 1971: 72-123.
[26] Tanaka R. Types of resting nuclei in Orchidaceae[J]. Shokubutsugaku Zasshi,1971,84 (993):118−122. doi: 10.15281/jplantres1887.84.118
[27] Ledingham GF. Chromosome numbers in Astragalus and Oxytropis[J]. Can J Genet Cytol,1960,2 (2):119−128. doi: 10.1139/g60-012
[28] 肖勇,杨耀东,夏薇,雷新涛,马子龙. 多倍体在植物进化中的意义[J]. 广东农业科学,2013,40(16):127−130. Xiao Y,Yang YD,Xia W,Lei XT,Ma ZL. Significance of polyploidy evolutionary in flowering plants[J]. Guangdong Agricultural Sciences,2013,40 (16):127−130. Xiao Y, Yang YD, Xia W, Lei XT, Ma ZL. Significance of polyploidy evolutionary in flowering plants[J]. Guangdong Agricultural Sciences, 2013, 40(16): 127-130.
[29] De Storme N,Geelen D. The impact of environmental stress on male reproductive development in plants:biological processes and molecular mechanisms[J]. Plant Cell Environ,2014,37:1−18. doi: 10.1111/pce.12142
[30] Van de Peer Y,Ashman TL,Soltis PS,Soltis DE. Polyploidy:an evolutionary and ecological force in stressful times[J]. Plant Cell,2021,33:11−26. doi: 10.1093/plcell/koaa015
[31] 刘玉红. 五种黄芪属植物的核型分析[J]. 植物分类学报,1984,22(2):125−127. Liu YH. Karyotype analysis of 5 species of genus Astragalus[J]. Journal of Systematics and Evolution,1984,22 (2):125−127. Liu YH. Karyotype analysis of 5 species of genus Astragalus[J]. Journal of Systematics and Evolution, 1984, 22(2): 125-127.
-
期刊类型引用(7)
1. 李珂佳,蔡晨,阮广鸣,凌书伟,潘标志,周家贵,刘金福,郑世群. 小叶红豆群落主要树种的种间关系及群落稳定性. 应用生态学报. 2025(02): 427-436 .  百度学术
百度学术
2. 谢婉丽,王奇悦,王秋雪,甘婉怡,武艳芳,黄柳菁. 闽江福州段自生草本植物生态位和种间联结研究. 热带亚热带植物学报. 2025(02): 149-158 . 百度学术
3. 商乃演,李东海,杨小波,黄耀,刘人通,苏欣,杜春雁,和绍翠,祁天运. 濒危植物蝴蝶树所在群落不同林层优势树种生态位与种间联结. 生态学报. 2025(07): 3389-3400 . 百度学术
4. 叶家桐,邓涛,岑华飞,段云博,朱晓珍,胡兴华. 广西银竹老山资源冷杉群落乔木优势树种生态位与种间联结. 生态学报. 2025(08): 3921-3932 . 百度学术
5. 向盈盈,唐绍清,卢永彬. 基于SSR标记淡黄金花茶的遗传多样性和遗传结构研究. 广西植物. 2025(04): 654-666 . 百度学术
6. 覃智敏. 不同种植密度对金花茶生长和开花的影响. 绿色科技. 2024(05): 179-182 . 百度学术
7. 艾鑫,薛卫星,艾训儒,姚兰,朱江,郭秋菊. 木林子矮曲林优势种生态位与种间联结性. 森林与环境学报. 2024(05): 501-510 . 百度学术
其他类型引用(2)

































计量
- 文章访问数: 213
- HTML全文浏览量: 79
- PDF下载量: 30
- 被引次数: 9
