
Study on the clonal diversity and genetic structure of the Eichhornia crassipes (Mart.) Solms from the middle and lower reaches of the Yangtze River
-
摘要:
水流在水生入侵植物的传播过程中具有重要作用。凤眼蓝(Eichhornia crassipes (Mart.) Solms)是我国危害极大的水生入侵植物。本研究基于AFLP标记,对长江中下游14个凤眼蓝种群(共399个植株)进行了克隆多样性和遗传结构分析,以探究水流对凤眼蓝扩散的影响。结果显示,利用4对AFLP引物,共检测到186个位点,其中56个为多态性位点(占30.11%)。该区域凤眼蓝种群的克隆多样性(D=0.273)和遗传多样性(I=0.011,HE=0.017)均很低,是由于优势的克隆生长而导致。种群间遗传分化很小(GST=0.019);同时AMOVA分析结果表明,1.84%的遗传变异来自于种群间,98.16%来自于种群内部,且14个种群间存在1个共有基因型,说明水流是长江流域凤眼蓝传播的主要媒介。因而,控制水流将极大促进该物种的有效治理。
Abstract:Water flow plays a crucial role in the propagation of aquatic invasive plants. Eichhornia crassipes is a major invasive aquatic plant in China, causing significant harm. Using AFLP markers, this study explored the clonal diversity and genetic structure of 14 E. crassipes populations (comprising 399 individuals) from the middle and lower reaches of the Yangtze River. Based on four pairs of AFLP primers, 186 loci were identified throughout all populations, including 56 polymorphic loci (30.11%). The populations exhibited low clonal diversity (D = 0.273) and low genetic diversity (I = 0.011, HE = 0.017), due to the dominant clonal reproduction system of the species. Genetic differentiation among populations was minimal (GST = 0.019). Analysis of molecular variance (AMOVA) indicated that 1.84% of the genetic variation was attributed to differences among populations, while 98.16% originated from within populations. Additionally, one common genotype was shared by all 14 populations. This study highlighted the significant role of water flow as a primary medium for the dispersal of E. crassipes in the Yangtze River Basin. Therefore, controlling the dispersal of water flow would be a crucial measure for effectively managing the spread of this species.
-
水流通常是水生植物传播和分布的重要决定因素[1]。水生植物的种子、花粉及其断枝等繁殖体会随着水流传播[2],这将会有利于水生植物种群间的基因交流或快速占领新的生境。例如,中国西南地区濒危植物靖西海菜花(Ottelia acuminata (Gagnep.) Dandy )种群的传播过程中,水流起着关键的“廊道”作用[3]。同时水流也会导致外来入侵水生植物的恶意扩张,如大薸(Pistia stratiotes L.)通过匍匐枝的迅速克隆繁殖,在水流的介导下,入侵周围水域[4]。
凤眼蓝(Eichhornia crassipes (Mart.) Solms)原生于巴西,其蓝紫色花大而艳丽颇具有观赏价值,因而作为园艺植物被引入我国[5]。由于凤眼蓝的叶柄中空膨大,因此可以漂浮于水面,同时该物种具有强大的克隆繁殖能力。这些特征导致凤眼蓝能够在水流的作用下迅速占领不同河道或水域,造成当地水生生境中生物多样性剧烈下降,也会引发广泛的水道堵塞而严重阻碍河道运输和水利灌溉,成为主要的极具危害的入侵水生植物之一[6]。
在我国,凤眼蓝主要分布于长江流域和珠江流域[7]。长江流域是我国最大的冲积平原,是全球六大河流生态区生物多样性热点地区之一[8]。该区域数以千计的浅水湖泊历史上均与长江直接相连,近几十年该区域的多数湖泊修建大坝从而通过闸口与长江相连[9, 10]。在这种河湖一体的生态系统中,水流对水生植物的传播和种群维持具有主导作用[11],但也有一些水生植物由于堤坝闸口的建立等显示出较高的遗传分化[12, 13]。在这种情况下,长江水流对凤眼蓝在长江中下游地区的传播是否起到主导作用,长江流域不同湖泊中的凤眼蓝具有怎样的遗传格局?
了解外来入侵植物的传播途径和传播方式,是有效控制其恶性扩散的前提[14]。以往对凤眼蓝的遗传格局的研究均在全国范围内开展[15, 16],对于集中在一个流域内的研究尚未见报道。由于扩增片段长度多态性(Amplified Fragment Length Polymorphism,AFLP)具有灵敏度高、重复性好、多态性丰富等特点[17],本研究利用AFLP标记对长江中下游地区的14个凤眼蓝种群进行克隆多样性和遗传多样性评估,并分析水流在凤眼蓝的传播和分布过程中的作用,以期为凤眼蓝的有效治理提供遗传学基础数据。
1. 材料与方法
1.1 样本收集
2017年夏季,我们对长江中下游地区的凤眼蓝进行了广泛调查和采集,并把来自同一湖泊或同一段河流的个体认为是一个种群。采样时,为了尽量采集到不同克隆系的分株,两个体间的采样间隔大于10 m。最终,共收集到14个种群,每个种群22~35个体,总计399个凤眼蓝样本。新鲜健康的幼叶经清洗后放入硅胶中干燥保存,直至DNA提取(表1、图1)。
![]() 表 1 凤眼蓝种群的采样点、样本数量和生境Table 1. Location, habitat, and sample size of 14 Eichhornia crassipes populations
表 1 凤眼蓝种群的采样点、样本数量和生境Table 1. Location, habitat, and sample size of 14 Eichhornia crassipes populations种群名称
Population位置
Location经纬度
Latitude and longitude样本数量
Sample size生境
HabitatCH 长湖 30°24.37′ N,112°21.99′ E 29 湖泊 HH 洪湖 29°50.16′ N,113°18.60′ E 35 湖泊 HG 黄盖湖 29°41.05′ N,113°32.41′ E 32 湖泊 XL 西凉湖 29°55.61′ N,114°07.24′ E 32 湖泊 FT 斧头湖 29°58.11′ N,114°19.58′ E 27 湖泊 DH 东湖 30°32.87′ N,114°25.28′ E 25 湖泊 ZDH 涨渡湖 30°37.31′ N,114°41.11′ E 26 湖泊 LG 龙感湖 29°58.21′ N,116°00.9′ E 31 湖泊 XH 西河 29°25.21′ N,116°38.32′ E 24 河流 MY 麻叶湖 29°18.72′ N,116°38.75′ E 28 湖泊 WC 武昌湖 30°16.69′ N,116°40.05′ E 30 湖泊 PG 破罡湖 30°35.72′ N,117°04.05′ E 32 湖泊 TH 太湖 31°15.21′ N,120°12.48′ E 22 湖泊 DS 淀山湖 31°11.63′ N,120°54.53′ E 26 湖泊 1.2 实验方法
1.2.1 DNA提取和PCR扩增
采用改进的CTAB法[18],每样本取约0.3 g干燥叶片用于提取总DNA。用1%的琼脂糖凝胶电泳检测DNA质量,用微量紫外分光光度计(ND2000)测定其浓度。将获得的高质量的DNA溶液浓度稀释为50 ng/µL,待用。
对已经合成的49对AFLP引物,进行扩增和筛选,筛选出4对条带清晰稳定的引物用于所有凤眼蓝样本的扩增(表2)。扩增程序主要包括:酶切、链接、预扩增和选扩增。预扩增反应体系为20 µL,包含4 µL限制性-连接酶,0.2 µL dNTPs,0.2 µL扩增引物和0.1 µL Taq DNA聚合酶(5 U),2 µL PCR缓冲液,13.3 µL ddH2O。预扩增程序为:94 ℃预热2 min;94 ℃ 20 s,56 ℃ 2 min,72 ℃ 2 min,共26个循环;60 ℃ 30 min结束。选择扩增的反应体系为20 µL,包含2 µL反应缓冲液,0.4 µL dNTPs(2 mmol/L),12.5 µL ddH2O,1 µL扩增引物(25 ng),0.1 µL Taq DNA聚合酶(5 U)和3 µL稀释后的预扩增模板。选择扩增程序为:94 ℃预变性2min;9个如下循环: 94 ℃ 20 s,66 ℃ 30 s,72 ℃ 2 min,每循环温度降低1 ℃;20个如下循环:94 ℃ 20 s,56 ℃ 30 s,72 ℃ 2 min;最后60℃保持30 min结束程序。
表 2 凤眼蓝种群AFLP分析中使用的接头、引物序列和不同引物组合的扩增多态性Table 2. Adaptor and primer pair sequences and number of total and polymorphic bands in AFLP analysis of Eichhornia crassipes序列类别
Sequence properties序列编号
Sequence code序列组成
Components of sequence总条带
Total bands多态性条带
Polymorphic bands多态性
Polymorphism / %接头序列 MseⅠ adaptor 5′ -GACGATGAGTCCTGAG-3′
5′-TACTCAGGACTCAT-3′EcoRⅠ adaptor 5′-CTCGTAGACTGCGTACC-3′
5′-AATTGGTACGCAGTCTAC-3′预扩增引物 5′-GTAGACTGCGTACCAATTCA-3′
5′-GACGATGAGTCCTGAGTAAC-3′选择扩增引物组合 E-AGC/M-CTG 45 17 37.78 E-AAG/M-CTC 46 13 28.26 E-AAC/M-CTG 48 14 29.17 E-AGA/M-CTC 47 12 25.53 选择扩增产物经6%变性聚丙烯酰胺凝胶分离,将胶板进行银染显色,从而获得具有不同片段条带的胶板。
1.2.2 数据分析
以25 bp和50 bp的Marker作为参照,人工读取50~800 bp的条带。对于同一等位基因位点(即在同一水平线上),有条带则记为“1”,无条带则记为“0”,最后形成由“1”和“0”组成的二维数据矩阵。
运用Genodive / GenoType软件[19]计算基因型(基株)总数(G)、Simpson多样性指数(D)、Fager指数(E)。采用POPGENE32[20]软件计算种群的多态性位点百分比(PP)、Nei’s遗传多样性(HE)[21]、Shannon’s指数(I)[22]和遗传分化指数(GST)。种群间基因流(每代迁移数)Nm用如下公式计算:Nm = (1- GST)/4GST [23] 。
用GenAIEx6.5 [24]软件的Mantel检验检测遗传距离与地理距离之间是否相关,同时使用该软件进行AMOVA分析。使用NTSYS软件[25],依据Nei’s [21]无偏遗传距离,对14个凤眼蓝种群进行UPGMA(非加权组平均法)聚类分析。
2. 结果与分析
2.1 凤眼蓝种群的克隆多样性
本研究使用4对引物共产生了186个位点,其中56个具有多态性(30.11%)。每个种群的克隆基因型(基株)数(G)为2~8,所有种群都是由多基因型组成,没有发现单克隆种群。凤眼蓝种群的平均克隆大小(NC)为7.764,变动范围为2.750(TH)~17.200(HH)(表3)。
表 3 14个凤眼蓝种群的克隆多样性Table 3. Clonal diversity of 14 Eichhornia crassipes populations种群Population 样本数N 基株数G 基因型比率G/N Simpson多样性指数D 平均克隆大小NC Fager指数E CH 29 7 0.241 0.429 4.140 0.244 HH 35 2 0.057 0.057 17.200 0.529 HG 32 4 0.125 0.236 8.000 0.324 XL 32 2 0.063 0.063 16.000 0.532 FT 27 6 0.222 0.342 4.500 0.249 DH 25 3 0.120 0.157 8.330 0.392 ZDH 26 3 0.115 0.151 8.670 0.390 LG 31 4 0.129 0.385 7.750 0.399 XH 24 2 0.083 0.083 12.000 0.543 MY 28 7 0.250 0.442 4.000 0.249 WC 30 7 0.233 0.366 4.290 0.221 PG 32 7 0.219 0.345 4.570 0.215 TH 22 8 0.364 0.546 2.750 0.261 DS 26 4 0.154 0.222 6.500 0.318 平均值 - 4.714 0.170 0.273 7.764 0.348 注:名称缩写同表1。下同。 Note: Abbreviations are the same as those in Table 1. Same below. 399个凤眼蓝样本共检测到66个基因型,14个种群间存在1个共有的基因型,另有3个基因型分别分布在两个湖泊中(附表1
1 ))。Simpson指数(D)平均为0.273,其变动范围为0.057(HH)~0.546(TH)。Fager指数(E)平均为0.348,在0.215(PG)~0.543(XH)之间变动。基因型比率(G/N)平均为0.170。2.2 凤眼蓝种群的遗传多样性
在种群水平上(表4),多态性位点百分比(Pp)为0.54%(XL)~9.14%(TH),平均值为3.46%;Nei's遗传多样性(HE)平均值为0.011,范围为0.002(XL)~0.032(TH);Shannon’s指数(I)平均值为0.017,范围为0.003(XL)~0.049(TH),种群间基因流Nm为12.908,说明不同种群之间的基因交流频繁。
表 4 14个凤眼蓝种群的遗传多样性Table 4. Genetic diversity of 14 Eichhornia crassipes populations种群
Population基因型数量
G多态性位点百分比
PP / %Nei's 遗传多样性
HEShannon's指数
ICH 7 3.23 0.010 (0.056) 0.016 (0.088) HH 2 1.08 0.003 (0.029) 0.005 (0.047) HG 4 2.15 0.007 (0.049) 0.011 (0.075) XL 2 0.54 0.002 (0.021) 0.003 (0.034) FT 6 8.06 0.026 (0.087) 0.040 (0.136) DH 3 1.08 0.003 (0.033) 0.005 (0.052) ZDH 3 1.08 0.003 (0.033) 0.005 (0.050) LG 4 2.15 0.007 (0.052) 0.011 (0.078) XH 2 1.61 0.005 (0.041) 0.008 (0.064) MY 7 8.06 0.023 (0.088) 0.040 (0.137) WC 7 3.23 0.011 (0.058) 0.016 (0.090) PG 7 3.23 0.010 (0.054) 0.016 (0.086) TH 8 9.14 0.032 (0.102) 0.049 (0.155) DS 4 3.76 0.012 (0.063) 0.019 (0.097) 平均值 4.7 3.46 0.011 (0.001) 0.017 (0.002) 物种水平 − 30.11 0.003 (0.006) 0.009 (0.017) 注:括号内容为标准误差。 Note: Standard errors are in parenthese. 2.3 遗传结构
本研究中发现凤眼蓝种群间遗传分化很小(GST=0.019)。AMOVA分析显示,1.84%的遗传变异来自于种群之间,而98.16%的遗传变异来自于种群内部(表5)。种群间的遗传距离与地理距离无相关性(r=0.082,P=0.240)。
表 5 凤眼蓝14个种群399个个体的AMOVA分析结果Table 5. Analysis of molecular variance (AMOVA) for 399 individuals among and within 14 natural Eichhornia crassipes populations变异来源
Source of variation自由度
df.平方和
Sum of squares方差分量
Variance components方差分量百分率
Percentage of variation / %显著性
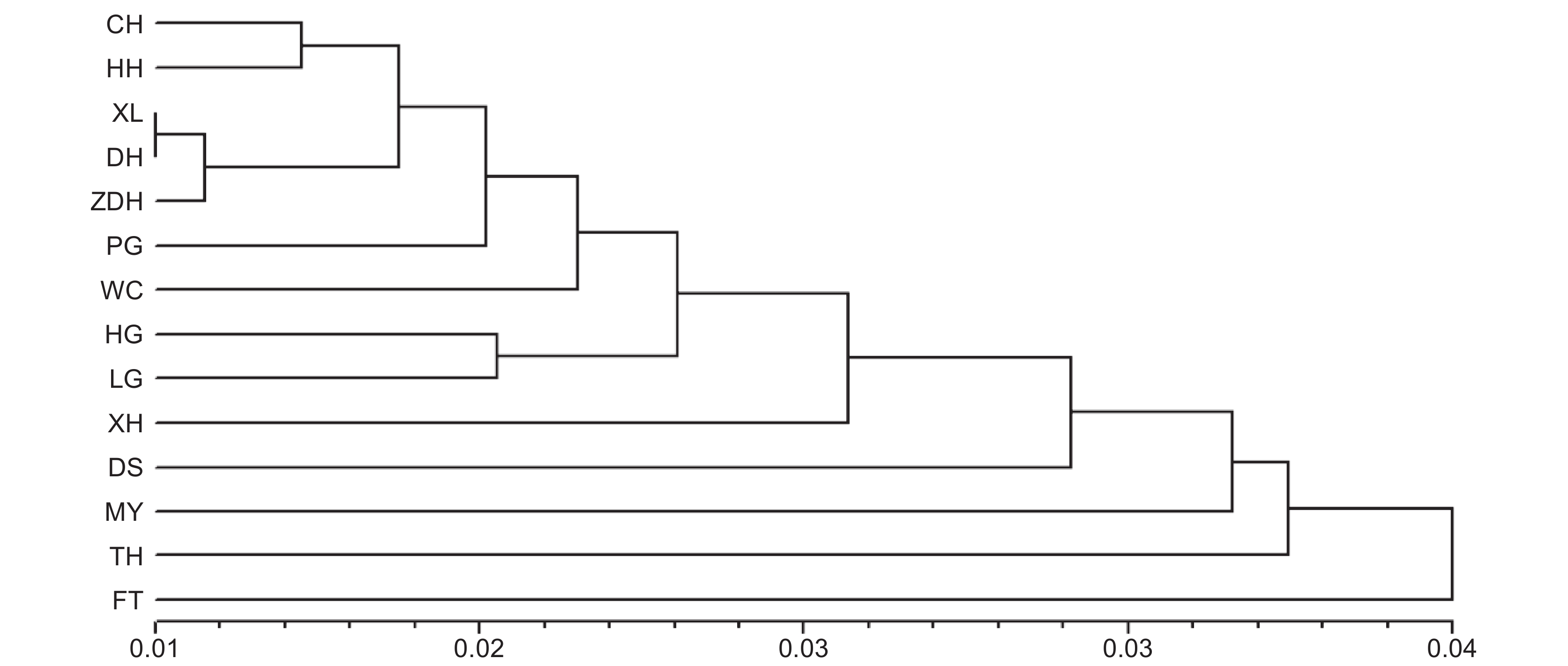
P种群间 13 5.250 0.005 1.84 <0.001 种群内 385 102.906 0.267 98.16 <0.001 UPGMA聚类中,少数地理距离较近的湖泊聚在一起,如长湖(CH)和洪湖(HH),西凉湖(XL)、东湖(DH)和涨渡湖(ZDH)。但是多数湖泊并未按照地理距离聚类。
3. 讨论
3.1 克隆多样性
本研究中,长江中下游的14个凤眼蓝种群显示出低的克隆多样性(Simpson多样性指数D=0.273),远低于Ellstrand和Roose[16]对21种克隆植物总结出来的等位酶克隆多样性(D=0.62)和Widén等[26]对45种克隆植物总结的RAPD克隆多样性(D=0.75)。本研究结果表明凤眼蓝依赖于无性繁殖来繁衍后代并进行种群扩张,这在以往种群遗传研究中也有体现[15, 27, 28]。如Zhang等[28]对世界范围内54个凤眼蓝种群的1 140个分株进行AFLP分析,发现其中有41个单克隆种群,而中国的19个种群中仅有5个种群表现为多克隆。凤眼蓝种群中低的克隆多样性是由其强大的克隆能力及有限的有性生殖能力而导致。凤眼蓝植株依靠其匍匐茎进行克隆生长,每个基株可以有10个以上的分株或次级分株[29],这也在我们的调查中被直接观察到。由于凤眼蓝在进行有性生殖时,其胚珠很容易败育导致种子产量较低,并且其种子成熟后落于深水区会导致萌发失败[29]。本研究的14个种群均来自于长江中下游湖区或通江河流中,水深较深且水面凤眼蓝长成厚重的“浮垫”,其水下的种子在这种情况下将很难萌发,我们亦在整个调查过程中未发现实生苗。
本研究中表示克隆均匀度的Fager指数(E)平均为0.348,低于Ellstrand和Roose[16]总结的结果(E=0.68),说明其种群内基因型分布非常不均匀。此外,凤眼蓝的克隆大小(Nc)在不同种群之间具有明显的差异(2.750~17.200),这反映了不同克隆的生长速度、繁殖能力以及适应不同环境条件的能力。值得注意的是,本研究中发现1个基因型在所有种群中共有,另外有3个基因型分别共享于两个种群中,这直接显示了水流对传播凤眼蓝的克隆分株起到至关重要的作用。
3.2 遗传多样性和遗传结构
遗传多样性高低反映物种对环境的适应能力,遗传多样性越高越能适应多变的环境 [30]。本研究结果表明,长江中下游地区的14个凤眼蓝种群表现出较低的遗传多样性水平(I=0.011,HE=0.017),这与Ren等[31]利用RAPD标记分析的中国凤眼蓝种群获得较低的遗传多样性结果相符(HE=0.046),而对于以往研究中单克隆的凤眼蓝种群则无法考察到其遗传多样性[15, 28]。有研究表明,克隆生殖的植物遗传多样性会比较低,原因是无性生殖方式缺乏有性生殖过程中染色体重组等,产生的克隆分株和基株的基因型完全一致[32-34]。因而,长江中下游的凤眼蓝种群由于其优势的克隆繁殖而导致其种群内遗传多样性过低。
水流连通性对于水生植物的遗传结构通常起到主导作用[35]。本研究中,来自于长江中下游的14个凤眼蓝种群显示了很微弱的遗传分化(GST=0.019),这和AMOVA分析的结果一致,该分析显示种群之间仅有1.84%的分化。这种微弱的遗传结构是由于种群之间频繁的基因流而导致(Nm=12.908)。考虑到凤眼蓝的植物种群扩张主要依靠传播克隆分株且本次研究范围跨度1 000多公里,频繁的基因流也印证了水流对其分株有极大的传播能力。
有趣的是,本研究中UPGMA聚类分析以及Mantel检验并未显示来源于相邻湖泊或河流的种群遗传关系更近。这可能是由于近几十年长江流域的多数湖泊河流都已修建水闸,仅在洪汛期或者枯水期闸门会因调节水位而被打开。可以推测,提供克隆分株的湖泊/河流要和接受的湖泊/河流在近乎同一时期打开闸门才可以完成迁移,而水位调节是受地方政府控制。因而,凤眼蓝分株未必会优先扩散到临近的湖泊。本研究中,除了1个被所有种群共有的基因型外,还有3个基因型分别被两个湖泊所共有,然而共有的湖泊并不邻近,这一结果也验证了以上的推论。
3.3 关于管理策略的建议
凤眼蓝于1901年引入我国,在南方各省作为观赏花卉或动物饲料被推广种植,其迅速占领长江及长江以南众多水域,对当地水生植物多样性及航运灌溉等都造成极大危害[36, 37]。本研究对长江中下游区域凤眼蓝种群的克隆及遗传结构进行研究,其结果对于该区域凤眼蓝的有效防治具有重要意义。
本研究发现凤眼蓝在该区域克隆多样性低且遗传分化小,造成其在该区域湖泊和河流中肆虐的主要原因是该物种本身强大的克隆生长能力,以及该区域内由长江连接的‘河湖一体’的水网结构。凤眼蓝植株的克隆繁殖能力使其不需要依赖周期漫长的有性繁殖来更新后代,因而繁殖更为便捷,其分株通过连接的水体散布到新的分布区,并快速扩散繁殖,占领水面。基于此,我们提出两点管理建议:(1)定期打捞水体中的凤眼蓝,尤其是与长江相连的各个闸口及河道内的凤眼蓝,这样可一定程度上阻断不同种群间克隆个体的散布;(2)对于未有凤眼蓝的水体,要加强巡查,一旦发现少数个体,就应迅速清理,因为考虑到其强大的克隆能力,不及时清除很易引起泛滥。
1 如需查阅附表内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
表 1 凤眼蓝种群的采样点、样本数量和生境
Table 1 Location, habitat, and sample size of 14 Eichhornia crassipes populations
种群名称
Population位置
Location经纬度
Latitude and longitude样本数量
Sample size生境
HabitatCH 长湖 30°24.37′ N,112°21.99′ E 29 湖泊 HH 洪湖 29°50.16′ N,113°18.60′ E 35 湖泊 HG 黄盖湖 29°41.05′ N,113°32.41′ E 32 湖泊 XL 西凉湖 29°55.61′ N,114°07.24′ E 32 湖泊 FT 斧头湖 29°58.11′ N,114°19.58′ E 27 湖泊 DH 东湖 30°32.87′ N,114°25.28′ E 25 湖泊 ZDH 涨渡湖 30°37.31′ N,114°41.11′ E 26 湖泊 LG 龙感湖 29°58.21′ N,116°00.9′ E 31 湖泊 XH 西河 29°25.21′ N,116°38.32′ E 24 河流 MY 麻叶湖 29°18.72′ N,116°38.75′ E 28 湖泊 WC 武昌湖 30°16.69′ N,116°40.05′ E 30 湖泊 PG 破罡湖 30°35.72′ N,117°04.05′ E 32 湖泊 TH 太湖 31°15.21′ N,120°12.48′ E 22 湖泊 DS 淀山湖 31°11.63′ N,120°54.53′ E 26 湖泊  下载: 导出CSV
下载: 导出CSV
表 2 凤眼蓝种群AFLP分析中使用的接头、引物序列和不同引物组合的扩增多态性
Table 2 Adaptor and primer pair sequences and number of total and polymorphic bands in AFLP analysis of Eichhornia crassipes
序列类别
Sequence properties序列编号
Sequence code序列组成
Components of sequence总条带
Total bands多态性条带
Polymorphic bands多态性
Polymorphism / %接头序列 MseⅠ adaptor 5′ -GACGATGAGTCCTGAG-3′
5′-TACTCAGGACTCAT-3′EcoRⅠ adaptor 5′-CTCGTAGACTGCGTACC-3′
5′-AATTGGTACGCAGTCTAC-3′预扩增引物 5′-GTAGACTGCGTACCAATTCA-3′
5′-GACGATGAGTCCTGAGTAAC-3′选择扩增引物组合 E-AGC/M-CTG 45 17 37.78 E-AAG/M-CTC 46 13 28.26 E-AAC/M-CTG 48 14 29.17 E-AGA/M-CTC 47 12 25.53
下载: 导出CSV
表 3 14个凤眼蓝种群的克隆多样性
Table 3 Clonal diversity of 14 Eichhornia crassipes populations
种群Population 样本数N 基株数G 基因型比率G/N Simpson多样性指数D 平均克隆大小NC Fager指数E CH 29 7 0.241 0.429 4.140 0.244 HH 35 2 0.057 0.057 17.200 0.529 HG 32 4 0.125 0.236 8.000 0.324 XL 32 2 0.063 0.063 16.000 0.532 FT 27 6 0.222 0.342 4.500 0.249 DH 25 3 0.120 0.157 8.330 0.392 ZDH 26 3 0.115 0.151 8.670 0.390 LG 31 4 0.129 0.385 7.750 0.399 XH 24 2 0.083 0.083 12.000 0.543 MY 28 7 0.250 0.442 4.000 0.249 WC 30 7 0.233 0.366 4.290 0.221 PG 32 7 0.219 0.345 4.570 0.215 TH 22 8 0.364 0.546 2.750 0.261 DS 26 4 0.154 0.222 6.500 0.318 平均值 - 4.714 0.170 0.273 7.764 0.348 注:名称缩写同表1。下同。 Note: Abbreviations are the same as those in Table 1. Same below.
下载: 导出CSV
表 4 14个凤眼蓝种群的遗传多样性
Table 4 Genetic diversity of 14 Eichhornia crassipes populations
种群
Population基因型数量
G多态性位点百分比
PP / %Nei's 遗传多样性
HEShannon's指数
ICH 7 3.23 0.010 (0.056) 0.016 (0.088) HH 2 1.08 0.003 (0.029) 0.005 (0.047) HG 4 2.15 0.007 (0.049) 0.011 (0.075) XL 2 0.54 0.002 (0.021) 0.003 (0.034) FT 6 8.06 0.026 (0.087) 0.040 (0.136) DH 3 1.08 0.003 (0.033) 0.005 (0.052) ZDH 3 1.08 0.003 (0.033) 0.005 (0.050) LG 4 2.15 0.007 (0.052) 0.011 (0.078) XH 2 1.61 0.005 (0.041) 0.008 (0.064) MY 7 8.06 0.023 (0.088) 0.040 (0.137) WC 7 3.23 0.011 (0.058) 0.016 (0.090) PG 7 3.23 0.010 (0.054) 0.016 (0.086) TH 8 9.14 0.032 (0.102) 0.049 (0.155) DS 4 3.76 0.012 (0.063) 0.019 (0.097) 平均值 4.7 3.46 0.011 (0.001) 0.017 (0.002) 物种水平 − 30.11 0.003 (0.006) 0.009 (0.017) 注:括号内容为标准误差。 Note: Standard errors are in parenthese.
下载: 导出CSV
表 5 凤眼蓝14个种群399个个体的AMOVA分析结果
Table 5 Analysis of molecular variance (AMOVA) for 399 individuals among and within 14 natural Eichhornia crassipes populations
变异来源
Source of variation自由度
df.平方和
Sum of squares方差分量
Variance components方差分量百分率
Percentage of variation / %显著性
P种群间 13 5.250 0.005 1.84 <0.001 种群内 385 102.906 0.267 98.16 <0.001
下载: 导出CSV
-
[1] O'Hare MT. Aquatic vegetation:a primer for hydrodynamic specialists[J]. J Hydraul Res,2015,53(6):687−698. doi: 10.1080/00221686.2015.1090493
[2] 董旭辉,张清慧,姚敏,葛亚汶,张永东,羊向东. 基于湖泊沉积物多指标的水生植被长期演替过程的定量重建[J]. 第四纪研究,2018,38(4):996−1006. doi: 10.11928/j.issn.1001-7410.2018.04.17 Dong XH,Zhang QH,Yao M,Ge YW,Zhang YD,Yang XD. Quantitative reconstruction of historical aquatic plant evolution based on multi-proxy analysis on lake sediment[J]. Quaternary Sciences,2018,38(4):996−1006. doi: 10.11928/j.issn.1001-7410.2018.04.17
[3] Fan XR,Njeri HK,Li W,Chen YY. Abundant historical gene flow within and among river systems for populations of Ottelia acuminata var. jingxiensis,an endangered macrophyte from Southwest China[J]. Aquat Bot,2019,157:1−9. doi: 10.1016/j.aquabot.2019.05.002
[4] Qian ZH,Li Y,Yang JS,Shi T,Li ZZ,Chen JM. The chromosome‐level genome of a free-floating aquatic weed Pistia stratiotes provides insights into its rapid invasion[J]. Mol Ecol Resour,2022,22(7):2732−2743. doi: 10.1111/1755-0998.13653
[5] 叶森. 水生外来入侵植物遥感识别与预警研究——以凤眼莲为例[D]. 呼和浩特:内蒙古大学,2020:1−57. [6] 周晴,潘晓云. 中国南部基塘区农业模式的变迁与凤眼蓝的入侵[J]. 植物生态学报,2014,38(10):1093−1098. doi: 10.3724/SP.J.1258.2014.00103 Zhou Q,Pan XY. Changes of the dike-pond agricultural pattern and water hyacinth invasion in southern China[J]. Chinese Journal of Plant Ecology,2014,38(10):1093−1098. doi: 10.3724/SP.J.1258.2014.00103
[7] 秦智雅,陶景怡,胡辰,阮爱东. 我国水域水葫芦的分布·影响·防治措施[J]. 安徽农业科学,2016,44(28):81−84. doi: 10.3969/j.issn.0517-6611.2016.28.026 Qin ZY,Tao JY,Hu C,Ruan AD. Distribution,influence and control measures of Eichhornia crassipes in China[J]. Journal of Anhui Agricultural Sciences,2016,44(28):81−84. doi: 10.3969/j.issn.0517-6611.2016.28.026
[8] Cao Y,Fan XR,Njeri HK,Pu YH,Li W,Chen YY. The correlation between genotype richness of submerged macrophytes and periphyton biomass:a mesocosm study based on five dominant submerged macrophytes from Yangtze River[J]. Plants,2023,12(13):2492. doi: 10.3390/plants12132492
[9] 周冬妮,尚钦. 河湖连通在长江中下游湖泊水环境治理与生态保护中的作用分析[C]//健康湖泊与美丽中国——第三届中国湖泊论坛暨第七届湖北科技论坛论文集. 武汉:中国科学技术协会,湖北省人民政府,2013:518−522. [10] 胡春宏,张双虎. 长江经济带水安全保障与水生态修复策略研究[J]. 中国工程科学,2022,24(1):166−175. doi: 10.15302/J-SSCAE-2022.01.018 Hu CH,Zhang SH. Strategies for water security and aquatic ecosystem restoration in the Yangtze River economic belt[J]. Strategic Study of CAE,2022,24(1):166−175. doi: 10.15302/J-SSCAE-2022.01.018
[11] Merritt DM,Nilsson C,Jansson R. Consequences of propagule dispersal and river fragmentation for riparian plant community diversity and turnover[J]. Ecol Monogr,2010,80(4):609−626. doi: 10.1890/09-1533.1
[12] Chen YY,Li XL,Yin LY,Li W. Genetic diversity of the threatened aquatic plant Ottelia alismoides in the Yangtze River[J]. Aquat Bot,2008,88(1):10−16. doi: 10.1016/j.aquabot.2007.08.002
[13] Han YC,Teng CZ,Zhong S,Zhou MQ,Hu ZL,Song YC. Genetic variation and clonal diversity in populations of Nelumbo nucifera (Nelumbonaceae) in central China detected by ISSR markers[J]. Aquat Bot,2007,86(1):69−75. doi: 10.1016/j.aquabot.2006.09.007
[14] 杜云安,杨连勇,杜华安,韦玮,唐小美,等. 洞庭湖区外来入侵植物调查分析及防治对策[J]. 生物安全学报,2020,29(3):222−228. Du YA,Yang LY,Du HA,Wei W,Tang XM,et al. An investigation of alien invasive plants around Dongting Lake area and their control countermeasures[J]. Journal of Biosafety,2020,29(3):222−228.
[15] Li WG,Wang BR,Wang JB. Lack of genetic variation of an invasive clonal plant Eichhornia crassipes in China revealed by RAPD and ISSR markers[J]. Aquat Bot,2006,84(2):176−180. doi: 10.1016/j.aquabot.2005.09.008
[16] Ellstrand NC,Roose ML. Patterns of genotypic diversity in clonal plant species[J]. Am J Bot,1987,74(1):123−131. doi: 10.1002/j.1537-2197.1987.tb08586.x
[17] 马庆国,王治军,徐慧敏,李明昆,李慧民,等. 金丝楸优良无性系遗传多样性和亲缘关系的AFLP分析[J]. 林业科学研究,2020,33(5):145−153. Ma QG,Wang ZJ,Xu HM,Li MK,Li HM,et al. AFLP analysis of Catalpa bungei superior clones[J]. Forest Research,2020,33(5):145−153.
[18] Doyle JJ,Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J]. Phytochem Bull,1987,19(1):11−15.
[19] Meirmans PG,van Tienderen PH. GENOTYPE and GENODIVE:two programs for the analysis of genetic diversity of asexual organisms[J]. Mol Ecol Notes,2004,4(4):792−794. doi: 10.1111/j.1471-8286.2004.00770.x
[20] Yeh FC,Yang R,Boyle TJ,Ye Z,Xiyan J,Yang R. PopGene 32,Microsoft windows-based freeware for population genetic analysis,Version 1.32[J]. Mol Biol Biotechnol Cent,Univ Alberta,Edmonton,2000:1−31.
[21] Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals[J]. Genetics,1978,89(3):583−590. doi: 10.1093/genetics/89.3.583
[22] Shannon CE,Weaver W. The Mathematical Theory of Communication[M]. Urbana:University of Illinois Press,1949,60(4):144.
[23] Slatkin M,Barton NH. A comparison of three indirect methods for estimating average levels of gene flow[J]. Evolution,1989,43(7):1349−1368. doi: 10.2307/2409452
[24] Peakall R,Smouse PE. GENALEX 6:genetic analysis in Excel. Population genetic software for teaching and research[J]. Mol Ecol Notes,2006,6(1):288−295. doi: 10.1111/j.1471-8286.2005.01155.x
[25] Rohlf FJ. NTSYS-pc:Numerical taxonomy and multivariate analysis system[J]. Exeter Software,Setauket,New York 1990.
[26] Widén B,Cronberg N,Widén M. Genotypic diversity,molecular markers and spatial distribution of genets in clonal plants,a literature survey[J]. Folia Geobot,1994,29(2):245−263. doi: 10.1007/BF02803799
[27] Ren MX,Zhang QG. Clonal diversity and structure of the invasive aquatic plant Eichhornia crassipes in China[J]. Aquat Bot,2007,87(3):242−246. doi: 10.1016/j.aquabot.2007.06.002
[28] Zhang YY,Zhang DY,Barrett SCH. Genetic uniformity characterizes the invasive spread of water hyacinth (Eichhornia crassipes),a clonal aquatic plant[J]. Mol Ecol,2010,19(9):1774−1786. doi: 10.1111/j.1365-294X.2010.04609.x
[29] 任明迅,张全国,张大勇. 入侵植物凤眼蓝繁育系统在中国境内的地理变异[J]. 植物生态学报,2004,28(6):753−760. doi: 10.3321/j.issn:1005-264X.2004.06.002 Ren MX,Zhang QG,Zhang DY. Geographical variation in the breeding systems of an invasive plant,Eichhornia crassipes,within China[J]. Acta Phytoecologica Sinica,2004,28(6):753−760. doi: 10.3321/j.issn:1005-264X.2004.06.002
[30] 秦惠珍,盘波,赵健,邹蓉,韦霄,唐凤鸾. 极小种群野生植物白花兜兰ISSR遗传多样性分析[J]. 广西科学,2022,29(6):1134−1140. Qin HZ,Pan B,Zhao J,Zou R,Wei X,Tang FL. Genetic diversity analysis by ISSR of Paphiopedilum emersonii,a plant species with extremely small populations[J]. Guangxi Sciences,2022,29(6):1134−1140.
[31] Ren MX,Zhang QG,Zhang DY. Random amplified polymorphic DNA markers reveal low genetic variation and a single dominant genotype in Eichhornia crassipes populations throughout China[J]. Weed Res,2005,45(3):236−244. doi: 10.1111/j.1365-3180.2005.00445.x
[32] Vallejo-Marín M,Dorken ME,Barrett SCH. The ecological and evolutionary consequences of clonality for plant mating[J]. Annu Rev Ecol Evol Syst,2010,41:193−213.
[33] Da Cunha NL,Xue HR,Wright SI,Barrett SCH. Genetic variation and clonal diversity in floating aquatic plants:comparative genomic analysis of water hyacinth species in their native range[J]. Mol Ecol,2022,31(20):5307−5325. doi: 10.1111/mec.16664
[34] Yu L,Stachowicz JJ,DuBois K,Reusch TBH. Detecting clonemate pairs in multicellular diploid clonal species based on a shared heterozygosity index[J]. Mol Ecol Res,2023,23(3):592−600. doi: 10.1111/1755-0998.13736
[35] Bornette G,Amoros C,Lamouroux N. Aquatic plant diversity in riverine wetlands:the role of connectivity[J]. Freshwater Biol,1998,39(2):267−283. doi: 10.1046/j.1365-2427.1998.00273.x
[36] 袁梨,吴登虎,姚勇,戴岗,田时飞,匡高翔. 水葫芦圈养对重庆动物园水禽湖富营养化水体水质的影响[J]. 养殖与饲料,2021,20(3):5−8. doi: 10.3969/j.issn.1671-427X.2021.03.003 [37] 程欣欣,聂丽云,余小玲,刘艳艳,马钰琦,等. 广西北部湾海岸带、海岛的外来入侵植物[J]. 热带亚热带植物学报,2023,31(3):444−454. doi: 10.11926/jtsb.4609 Cheng XX,Nie LY,Yu XL,Liu YY,Ma YQ,et al. Invasive alien plants in the coastal zone and islands of Beibu Gulf,Guangxi[J]. Journal of Tropical and Subtropical Botany,2023,31(3):444−454. doi: 10.11926/jtsb.4609
-
其他相关附件
-
DOCX格式
符丽妹附表1 点击下载(25KB)
-
计量
- 文章访问数: 148
- HTML全文浏览量: 27
- PDF下载量: 12
