
Protoraphis hustedtiana var. nana Takano, a newly recorded araphid diatom in China
-
摘要:
本研究报道了我国首次记录的一个无壳缝类硅藻变种——胡氏原缝藻微型变种(Protoraphis hustedtiana var. nana Takano)。该变种采自广西涠洲岛,为生活在马尾藻(Sargassum spp.)上的植表生硅藻(Epiphytic diatom)。利用光学显微镜和扫描电子显微镜对胡氏原缝藻微型变种的形态进行了详细的观察和描述,并介绍了该变种的生态习性和地理分布。
Abstract:This study reports a newly recorded araphid diatom, Protoraphis hustedtiana var. nana Takano, from Weizhou Island, China. This variety was found epiphytic on Sargassum spp. Its morphology was examined in detail using light and scanning electron microscopy, and its ecological habitat and geographical distribution were documented.
-
Keywords:
- Araphid diatom /
- Epiphytic diatom /
- New record /
- Variety /
- China
-
无壳缝类硅藻是一大类上、下壳面均不具有壳缝系统的硅藻类群,其种类繁多,目前已报道26属1 100余种,其中中国已记录23属220余种[1]。常见的无壳缝类硅藻属包括:缝舟藻属(Rhaphoneis)、斑条藻属(Grammatophora)、横纹藻属(Plagiostriata)、楔形藻属(Licmophora)和平片藻属(Tabularia)等[2-4]。此类硅藻可通过壳端分泌胶质并附着于水中各类基质表面,也有一些种类可在水体中以浮游形式生活[5]。根据金德祥先生[6]在1978年提出的硅藻分类系统,所有无壳缝类硅藻均属于等片藻目,但我国淡水硅藻学者常使用“无壳缝目”代替这一名称。然而,随着硅藻分类学的发展,无壳缝目硅藻所包含的种类发生了变化。例如,Round等[7]曾提出在硅藻门下设圆筛藻纲、脆杆藻纲和硅藻纲,其中,隶属于脆杆藻纲的无壳缝目泛指所有无壳缝类和拟壳缝类硅藻。
原缝藻属(Protoraphis)是一个典型的无壳缝类硅藻属[8]。目前,该属仅有2个种和1个变种:大西洋原缝藻(Protoraphis atlantica Gibson)、胡氏原缝藻(Protoraphis hustedtiana Simonsen)和胡氏原缝藻微型变种(Protoraphis hustedtiana var. nana Takano)[9-16]。本属细胞壳面整体呈披针形,顶端宽圆;线纹平行排列,由圆形、椭圆形或矩形的点纹组成;假壳缝狭长,呈明显的线形,且位于壳面正中间,但在靠近两端处向相反方向弯曲,并终止于两个椭圆形沟槽;端部沟槽贯穿壳面,具有增厚的硅质边缘,并在内部衍生出一系列特殊结构,这是本属的显著特征。假带状藻属(Pseudohimantidium)也具有相似的假壳缝和沟槽[17],虽然其壳面和顶纹区的形态特征与原缝藻属不同,但这两个属被合并到到原缝藻科中[12]。此外,原缝藻属与新针杆藻属(Neosynedra)、圆蕊藻属(Cyclophora)和肠形藻属(Lucanicum)在形态上也较为相似,它们均具有线形或披针形的壳面以及由若干狭缝组成的顶纹区,但通过光学和电子显微镜发现,这些硅藻属之间存在着明显的区分特征,如新针杆藻属在靠近壳端处的突起结构为内外两瓣组成的唇形突[18],圆蕊藻属在内壳面中部分布有个近圆形的假隔片[7],肠形藻属的线纹则由横向伸长的长室孔组成[19]。
作者于2021年4月对广西涠洲岛海域的植表生硅藻(Epiphytic diatoms)多样性进行调查时,在通过潜水采集的两种马尾藻(Sargassum spp.)上发现了一个我国首次记录的海洋硅藻变种——胡氏原缝藻微型变种(Protoraphis hustedtiana var. nana Takano)。本研究以金德祥分类系统为基础,结合光学显微镜和扫描电子显微镜的观察结果,对该变种的形态特征进行了详细描述,并讨论了该变种的生态分布特点,以期丰富我国海洋硅藻的研究内容和基础资料。
1. 材料与方法
2021年4月24日,本研究在对广西涠洲岛海域(21°1′6″ N,109°6′33″ E)大型海藻上的植表生硅藻多样性进行调查时,通过潜水采集了一批马尾藻样品,并对其体表的植表生硅藻进行了收集。涠洲岛是中国最大、地质年龄最年轻的火山岛,全岛面积约为24.74 km2,海岸线全场12.5 km。涠洲岛海域位于广西北海市南面,属热带季风气候,其海域及沿岸礁石上生长着大量马尾藻,具备丰富的海洋生物资源[20]。本研究中的硅藻标本,主要附着在涠洲马尾藻(S. weizhouense Tseng & Lu)和围氏马尾藻(S. withgii (Greville) J. Agardh)上。于实验室内,首先利用超声波细胞破碎仪对植表生硅藻进行了分离(300 W,25 s),并对样品使用5%的海水福尔马林溶液固定,保存于广西海洋科学院样品库中(样品编号:GXWZ5-1、GXWZ6-2);然后取5 mL样品于试管中,根据样品有机质含量按1∶1~1∶2不同体积比例加入浓硝酸,沸水浴中加热20 min,以去除样品中的有机质,待样品冷却,加满蒸馏水,自然沉降24 h,吸取上清液,再次加满蒸馏水,反复清洗样品直至样品溶液呈中性;最后吸取适量酸化后的硅藻样品,用Naphrax®或Mountmedia封片剂制成永久玻片,在Olympus CX33型光学显微镜下进行观察,并利用明美MSX2-H型显微摄影数码相机拍照,另过滤5 mL清洁后的硅藻样品于滤膜(孔径0.22 μm)上,待其自然风干后,随机剪取约5 mm×5 mm的小方块贴合到粘有导电胶的铝台上,经过喷金处理(30 mA,60 s)后,利用Hitachi S-3400N扫描电子显微镜进行观察和拍照。
本研究基于客观的观察结果,分别绘制了胡氏原缝藻微型变种的显微和超微结构形态图,结合所拍摄的照片,对该变种进行准确的形态学描述。
2. 结果与分析
胡氏原缝藻微型变种Protoraphis hustedtiana var. nana Takano.
Takano 1985, p. 32, figs 1–8; Sullivan 1993, p. 162, figs 1–8.
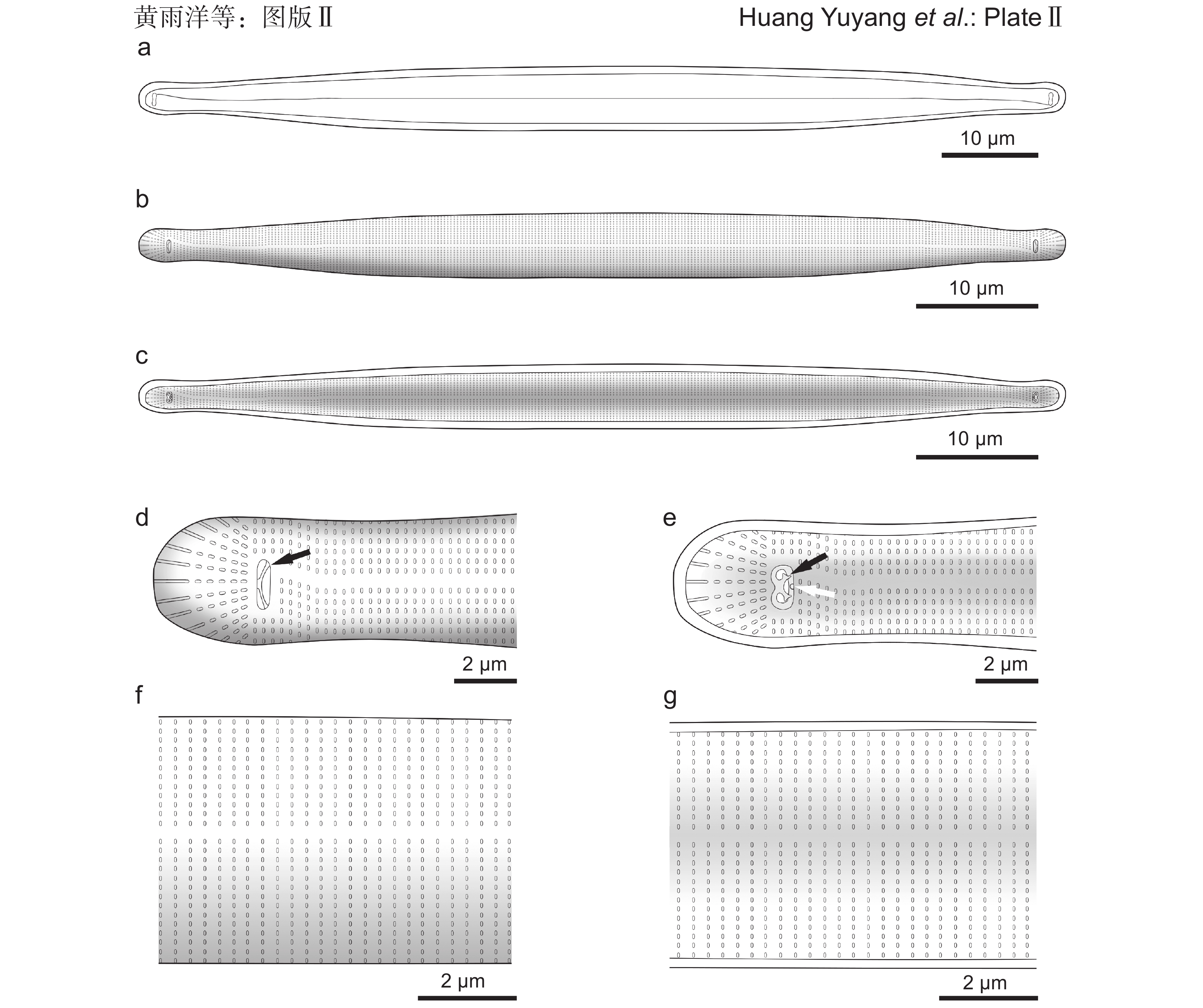
光学显微镜观察:壳面透明,呈披针形,两端宽圆且略成头状。长33~95 μm,宽4~7 μm(n=9)。假壳缝明显,呈线形,大部分位于壳面中间,但在靠近两端处向异侧弯曲,终止于两个近椭圆形的沟槽(图版Ⅰ:a、图版Ⅱ:a)。
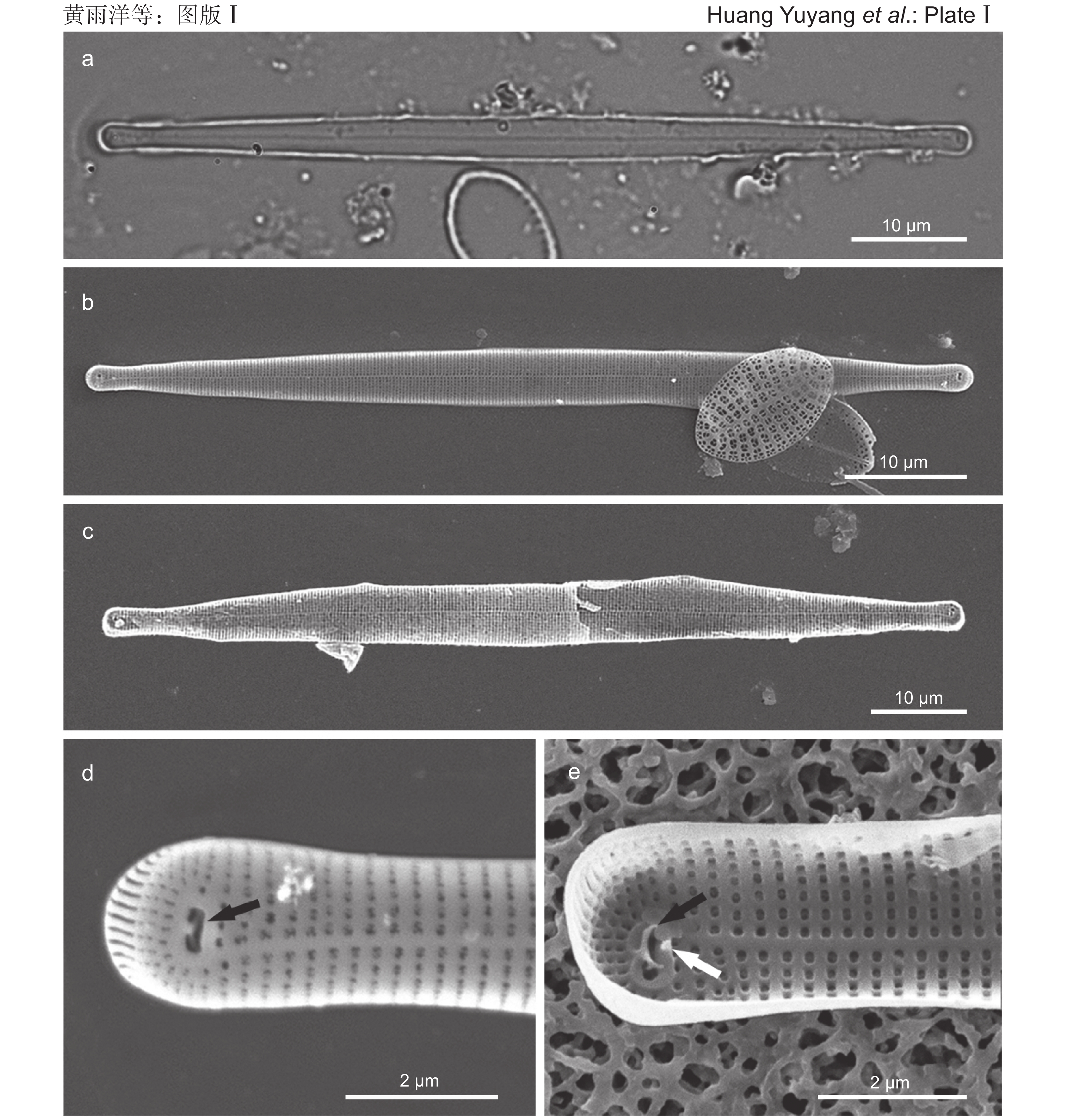
![]() 图版Ⅰ 胡氏原缝藻微型变种照片。a:光学显微镜下的形态;b~e:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺状或齿状突起)。图版Ⅰ. Micrographs of Protoraphis hustedtiana var. nana Takano.a: Light microscopy observation; b-e: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow).
图版Ⅰ 胡氏原缝藻微型变种照片。a:光学显微镜下的形态;b~e:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺状或齿状突起)。图版Ⅰ. Micrographs of Protoraphis hustedtiana var. nana Takano.a: Light microscopy observation; b-e: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow).![]() 图版Ⅱ 胡氏原缝藻微型变种手绘图。a:光学显微镜下的形态;b~g:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺或齿状突起);f:外壳面中部;g:内壳面中部。图版Ⅱ. Drawings of Protoraphis hustedtiana var. nana Takano.a: Light microscopy observation; b-g: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow); f: External view of mid-valve; g: Internal view of mid-valve.
图版Ⅱ 胡氏原缝藻微型变种手绘图。a:光学显微镜下的形态;b~g:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺或齿状突起);f:外壳面中部;g:内壳面中部。图版Ⅱ. Drawings of Protoraphis hustedtiana var. nana Takano.a: Light microscopy observation; b-g: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow); f: External view of mid-valve; g: Internal view of mid-valve.扫描电子显微镜观察结果显示,壳面呈棍棒状或披针形,对角对称,顶端宽圆(图版Ⅰ:b、图版Ⅱ:b)。外壳面,线纹由单行平行排列的椭圆形或矩形点纹组成,10 μm 内 32~37 条,横向点纹 1 μm 内 4~5 个(图版Ⅱ:f);假壳缝清晰显著,大部分居中且呈直线型,但在靠近两端处向异侧骤然弯曲并终止于沟槽,整体呈现出类似字母“S”的形态(图版Ⅰ:b、图版Ⅱ:b);沟槽为长椭圆形或矩形开口,长0.5~0.7 μm,横向分布,周围无点纹,贯穿硅质壳,其内具有一个弓形的硅质结构(图版Ⅰ:d ~ e、图版Ⅱ:d ~ e,黑色箭所示);壳端顶纹区由12~14条纵向的狭缝组成(图版Ⅰ:d、图版Ⅱ:d)。内壳面,沟槽的边缘增厚,形成耳状基部结构,其靠近壳端一侧衍生出1个“Y”形或鹿角状的硅质突起,另一侧则分布有1个发育不全的刺状或齿状突起(图版Ⅰ:e、图版Ⅱ:e,白色箭所示);线纹间的肋纹略发达(图版Ⅰ:e)。
生态习性:生活在海洋环境中,可附着在动物和植物体表。
地理分布:胡氏原缝藻微型变种首次记录于日本田边湾(Tanabe Bay)的一种螺类体表,其在美国密西西比湾(Missippi Sound)的海草床中也有分布。本研究中的标本采自广西涠洲岛(标本号:ED WZ5-6、ED WZ6-3),为中国首次报道。
3. 讨论
胡氏原缝藻微型变种的模式标本采自于日本田边湾[14],此后,该变种在美国密西西比湾也有所记录[16],其主要形态特征为靠近壳端处的沟槽及其衍生的 “Y”型硅质突起。本研究中采自广西涠洲岛的硅藻标本具备这一独特的结构,因此可以确定其为胡氏原缝藻微型变种。然而,涠洲岛种群与已报道的两个种群之间仍存在形态上的细微差异(表1):(1)在细胞大小方面,涠洲岛种群的细胞较大(长33~95 μm,宽4~7 μm),而模式种群和Sullivan[16]所报道的美国种群的细胞较小(前者长19~57 μm,宽4.5~5.8 μm;后者长32~48 μm,宽4.5 μm),尤其是最大个体的壳体长度上,涠洲岛种群比其他两个种群增长了近两倍;(2)在顶纹区组成方面,涠洲岛种群的狭缝数量最多达到了14条,而模式种群和Sullivan[16]所报道的美国种群分别仅有7条和12条;(3)在沟槽长度方面,涠洲岛种群呈现出相较于另外两个种群更短的沟槽,约为0.5 μm;(4)在线纹密度方面,涠洲岛种群的线纹密度范围更广(10 μm内32~37条),几乎涵盖了另两个种群所记录的数据范围(模式种群:10 μm内32条,Sullivan[16]所报道的美国种群:10 μm内34~36条)。
表 1 胡氏原缝藻微型变种不同种群之间的形态特征比较Table 1. Morphological characteristics of Protoraphis hustedtiana var. nana from different regions胡氏原缝藻微型变种与原缝藻属内其他物种在形态上的区别较为明显。首先,胡氏原缝藻微型变种在靠近壳端一侧具有一个“Y”形或鹿角状的硅质结构,另一侧有一个发育不全的刺状或齿状突起;胡氏原缝藻衍生出的硅质结构更为复杂,其靠近壳端一侧的硅质结构呈“E”形突起,而另一侧分布有一个薄片状或“U”形突起[21];大西洋原缝藻的突起则为齿状结构[10],该结构相较于属内其他物种更为简单。其次,与本研究所记录的胡氏原缝藻微型变种相比,Hallegraeff和McWilliam[15]所报道的胡氏原缝藻壳体更长(40~130 μm),而Gibson[10]所描述的大西洋原缝藻壳体更短(18~50 μm)。然后,胡氏原缝藻微型变种的点纹呈椭圆形或矩形,这与Gibson[10]所记录的大西洋原缝藻的点纹形状相似,却不同于胡氏原缝藻的圆形孔纹[21]。最后,本研究所测量的胡氏原缝藻微型变种沟槽长度为0.5~0.7 μm,远远短于胡氏原缝藻的沟槽长度(1.5 μm)[15]。
原缝藻属是一个寡种硅藻属,虽然其在自然界中相对罕见,但该属在海洋中有着较为广泛的分布。截至目前,原缝藻属硅藻在阿拉伯湾、波斯湾、美国、日本、澳大利亚、加勒比海、巴西和中国东海的海域中都有记录[21],本研究所描述的胡氏原缝藻微型变种采集于北部湾的涠洲岛周边海域,进一步扩大了其全球分布范围。原缝藻属的栖息环境较为特殊,它们一般附生于海洋动植物体表。据报道,大西洋原缝藻仅在海洋桡足类体表营动表生生活,其宿主包括幼平头水蚤(Candacia catula Giesbrecht)、短突简角水蚤(Pontellopsis brevis Giesbrecht)和简拟平头水蚤(Paracandacia simplex Giesbrecht)等 [11, 22];胡氏原缝藻也仅发现于大眼水蚤属(Corycaeus Dana)、异尾平头水蚤(Candacia discaudata A. Scott)和伯氏平头水蚤(C. bradyi A. Scott)等少数海洋桡足类的体表,且主要附着在海洋桡足类身体后附肢上,少数附着在身体的背部[15, 21]。然而,不同于大西洋原缝藻和胡氏原缝藻,胡氏原缝藻微型变种首次记录于日本田边湾里的一种螺类体表,其后发现于美国密西西比湾海草床中,本研究则在广西涠洲岛的马尾藻上发现了该变种。由此可见,胡氏原缝藻微型变种的宿主可能较为多样化,但该变种与其宿主之间的生态学模式尚不清晰,今后的研究可在此方面做进一步的了解。
-
![]()
图版Ⅰ 胡氏原缝藻微型变种照片。
a:光学显微镜下的形态;b~e:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺状或齿状突起)。
图版Ⅰ. Micrographs of Protoraphis hustedtiana var. nana Takano.
a: Light microscopy observation; b-e: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow).
![]()
图版Ⅱ 胡氏原缝藻微型变种手绘图。
a:光学显微镜下的形态;b~g:扫描电子显微镜下的形态。b:外壳面整体观;c:内壳面整体观;d:外壳面顶端(黑色箭所示为弓形硅质结构);e:内壳面顶端(黑色箭所示为弓形硅质结构,白色箭所示为发育不全的刺或齿状突起);f:外壳面中部;g:内壳面中部。
图版Ⅱ. Drawings of Protoraphis hustedtiana var. nana Takano.
a: Light microscopy observation; b-g: Scanning electron microscopy observation. b: External view of whole valve; c: Internal view of whole valve; d: External view of valve end (note siliceous structure indicated by black arrow); e: Internal view of valve end (note siliceous structure indicated by black arrow, and spine or tooth-like process indicated by white arrow); f: External view of mid-valve; g: Internal view of mid-valve.
表 1 胡氏原缝藻微型变种不同种群之间的形态特征比较
Table 1 Morphological characteristics of Protoraphis hustedtiana var. nana from different regions
 下载: 导出CSV
下载: 导出CSV
-
[1] 罗粉,尤庆敏,于潘,王全喜. 横断山区无壳缝目硅藻中国新记录[J]. 植物科学学报,2022,40(6):733−743. Luo F,You QM,Yu P,Wang QX. Newly recorded species of Araphidiales from the Hengduan Mountains[J]. Plant Science Journal,2022,40(6):733−743.
[2] 王震. 福建北部沿海沙生境无壳缝类硅藻的分类学研究[D]. 厦门:厦门大学,2020:1−10. [3] 金德祥,程兆第,林均民,刘师成. 中国海洋底栖硅藻类:上卷[M]. 北京:海洋出版社,1982:1−100. [4] 金德祥,程兆第,刘师成,马俊享. 中国海洋底栖硅藻类:下卷[M]. 北京:海洋出版社,1992:1−100. [5] 齐雨藻,李家英. 中国淡水藻志:第10卷[M]. 北京:科学出版社,2004:1−161. [6] 金德祥. 硅藻分类系统的探讨[J]. 厦门大学学报,1978(2):31−50. Chin TG. A discussion on the phylogenesis of diatoms[J]. Journal of Xiamen University,1978(2):31−50.
[7] Round FE,Crawford RM,Mann D G. The Diatoms:Biology & Morphology of the Genera[M]. Cambridge:Cambridge University Press,1990:345−447.
[8] Purushothaman A,Romagnoli T,Francis SV,Thomas LC,Padmakumar KB. First report of marine epizoic diatom,Protoraphis atlantica (Protoraphidaceae) on calanoid copepods along the southeastern Arabian Sea[J]. Symbiosis,2021,84(2):131−140. doi: 10.1007/s13199-021-00772-6
[9] Gibson RA. Pseudohimantidium pacificum,an epizoic diatom new to he Florida Current (Western North Atlantic Ocean)[J]. J Phycol,1978,14(3):371−373. doi: 10.1111/j.1529-8817.1978.tb00316.x
[10] Gibson RA. Protoraphis atlantica sp. nov.,a new marine epizoic diatom[J]. Bacillaria,1979,2:109−126.
[11] Gibson RA. An ultrastructure study of Pseudohimantidium pacificum Hust. & Krasske (Bacillariophyceae:Protoraphidaceae) with special reference to the labiate processes[J]. Nova Hedwigia Beiheft,1979,64:147−156.
[12] Simonsen R. Protoraphidaceae,eine neue Familie der Diatomeen[M]//Gerloff J,Cholnoky JB. Diatomaceae Ⅱ. Berlin:Beihefte zur Nova Hedwigia,1970:177−394.
[13] Simonsen R. The diatom plankton of the Indian Ocean expedition of R/V “Meteor” 1964-1965[J]. Meteor Forsch Ergeb Reihe D,1974,19:1−107.
[14] Takano H. A small form of the marine diatom Protoraphis hustediana,epizoic on a snail[J]. Bull Tokai Reg Fish Res Lab,1985,117:31−35.
[15] Hallegraeff GM,McWilliam PS. The complex labiate process of the epizoic diatom Protoraphis hustedtiana Simonsen[J]. Beihefte zur Nova Hedwigia,1990,100:39−45.
[16] Sullivan MJ. The labiate process of the diatom Protoraphis hustedtiana var. nana Takano[J]. Nova Hedwigia Beihefte,1993,106:161−167.
[17] Rivera PS,Gonzalez HE,Barrales HL. Cingulum and valve morphology of Pseudohimantidium Hustedt & Krasske (Bacillariophyceae)[J]. Phycologia,1986,25(1):19−27. doi: 10.2216/i0031-8884-25-1-19.1
[18] Williams DM,Round FE. Revision of the genus Synedra Ehrenb[J]. Diatom Res,1986,1(2):313−339. doi: 10.1080/0269249X.1986.9704976
[19] Lobban CS,Ashworth MP. Lucanicum concatenatum,gen. nov.,sp. nov.,a benthic marine diatom from Guam,and a less restrictive diagnosis for Cyclophorales (Bacillariophyta)[J]. Mar Biodivers Rec,2014,7:e90.
[20] 郭键林,孙显,杨宇峰,王庆. 广西涠洲岛大型海藻场及其邻近海域桡足类群落结构演替特征[J]. 热带海洋学报,2023,42(4):155−165. Guo JL,Sun X,Yang YF,Wang Q. The succession characteristics of copepod community structure in the seaweed bed and its adjacent waters in the Weizhou Island,Guangxi Province[J]. Journal of Tropical Oceanography,2023,42(4):155−165.
[21] Li L,Chen CP,Sun L,Zhang JW,Liang JR,Gao YH. Protoraphis Simonsen,a newly recorded marine epizoic diatom genus for China[J]. Acta Oceanol Sin,2020,39(4):120−126. doi: 10.1007/s13131-019-1467-z
[22] Gómez F,Wang L,Lin SJ. Morphology and molecular phylogeny of epizoic araphid diatoms on marine zooplankton,including Pseudofalcula hyalina gen. & comb. nov. (Fragilariophyceae,Bacillariophyta)[J]. J Phycol,2018,54(4):557−570. doi: 10.1111/jpy.12760
计量
- 文章访问数: 207
- HTML全文浏览量: 35
- PDF下载量: 29
