
Diversity, distribution patterns, and floristic characteristics of seed plants endemic to Tibet, China
-
摘要:
基于西藏特有种子植物名录,结合物种自然地理分布信息,对西藏特有种子植物科属种多样性组成特征、物种生活型多样性、垂直海拔分布及县域尺度分布格局、属水平植物区系性质进行了分析,并对特有种热点地区进行了识别。结果显示,西藏特有种子植物有1 079种,隶属89科297属,特有性高,其中含20种以上的科为19个,含特有种最多的科为菊科;含10种以上的属有24个,含特有种最多的属为翠雀属(Delphinium)和小檗属(Berberis)。多年生草本植物在特有种中占绝对优势(66.4%)。从区系成分来看,温带性质的属占主导(61.6%)。东南部的墨脱县含特有种数量最多(271种)。总体上,西藏特有种子植物的空间分布格局极不均匀,喜马拉雅东南麓是丰富度最高的区域。西藏特有种子植物的海拔分布范围宽泛,集中分布在3000 ~ 4500 m,特有种丰富度在海拔梯度上呈单峰分布格局。本研究结果可为将来确定西藏植物多样性保护优先次序、红色名录制定及自然保护体系的建设提供参考。
Abstract:Based on the list of endemic seed plants of Tibet, combined with the natural geographic distribution of species, the composition characteristics of family, genus, and species diversity, species life-form diversity, vertical altitude and county-scale distribution patterns, and floristic properties at the genus level were analyzed, with endemic species hotspots also identified. Results showed 1 079 species of endemic seed plants in Tibet, belonging to 89 families and 297 genera, with high specificity. Among the identified plants, 19 families contained more than 20 species, and Asteraceae contained the most endemic species. In total, 24 genera contained more than 10 species, with Delphinium and Berberis identified as the most endemic genera. Perennial herbs accounted for the highest proportion (66.4%) among endemic plants. In terms of floristic composition, temperate genera were dominant (61.6%). Medog County contained the largest number of endemic species (271 species). In general, the distribution pattern of endemic seed plants in Tibet was extremely uneven, with the southeast Himalayas showing the highest richness. The endemic seed plants also exhibited an extremely wide altitudinal range, showing a single-peak species-richness distribution pattern along the altitudinal gradient, concentrated at 3 000–4 500 m. The results of this study provide basic plant data to support the prioritization of plant diversity conservation in Tibet, the establishment of a red list, and the construction of a natural protection system in the future.
-
Keywords:
- Endemic species /
- Diversity disturbance patterns /
- Flora /
- Conservation /
- Tibet
-
生物类群的特有现象是生物地理学研究的重要组成部分[1]。随着物种灭绝的速度加快,特有物种在生物多样性保护研究中备受宏观生态学家与保护生物学家的关注[2]。特有种由于其狭域分布的特征,容易变成濒危物种,因此保护特有物种通常能更好地保护珍稀濒危物种[1, 2]。特有物种丰富度(Endemic species richness)是指局限分布于一定区域(行政区、自然保护区、某一山区等)的特有物种的数量。特有种是相对于一个特定的生物地理范围来说的,特有物种丰富度是确定生物多样性优先保护区的重要指标,对生物多样性科学理论研究与保护政策的制定意义重大[3-5]。另外,特有物种的地理空间分布格局也一直是生物学、生态学与保护生物学等众多领域关注的主题[5-8],这些研究的共同结论是,特有物种的空间分布不是随机的,它们在世界各地的分布极不均匀,往往集中在某一特定的区域或生境内。个别物种被发表后就再也没有采集到过其标本,甚至有些物种已濒危,但特有种对生态群落的发展与演化有着独特的作用[4, 6]。因此,评估特有物种空间分布格局对于制定有效的稀有物种保护策略、实施保护行动及界定其保护区范围是必要的。一个地区的特有程度被认为是衡量该地区生态系统独特性的重要指标,因此,特有种丰富度是确定生物多样性保护地点优先次序的最常用标准之一[9, 10]。目前,生物多样性面临急剧丧失的威胁,需要在全球、区域与地方各级中确定优先保护目标,以便将有限的资源集中在最重要的需求对象上[11, 12]。特有物种常常分布稀少,容易灭绝[4]。因此,研究特有物种是迫在眉睫的一项工作,目前为止,国内对某一特定区域特有物种的相关报道很多[13-18],为植物多样性保护与植物区系地理的研究奠定了良好的基础。
西藏高原是一个独特的自然地理单元,有着从热带到寒带所有垂直地带的植被带谱。复杂的地形地貌孕育了西藏丰富的植物区系多样性,特有种数量丰富[19]。但针对西藏特有种子植物层面进行多样性特征、区系性质及分布格局的研究报道还很鲜见。特有物种是一个地区重要的特色生物种质资源,有着极高的科学研究与保护价值,对特有植物的研究更有助于探讨西藏植物区系的发展历史与现状,也有利于特色植物资源的开发与利用。因此,本研究在收集整理西藏特有种子植物名录与物种分布数据的基础上,对科属种组成及其多样性特征、物种生活型多样性、物种垂直海拔分布格局及县域尺度(水平地理)分布格局、属水平植物区系性质等方面进行了分析研究。研究结果旨在回答:(1)目前被描述的西藏特有种子植物有多少种?其多样性组成结构如何及生活型组成比例是多少?(2)植物区系性质如何?(3)特有物种水平地理分布与海拔垂直分布的格局特征如何?研究结果对进一步了解西藏地区的植物多样性,特别是特有植物的基本情况具有重要意义,在一定程度上也可为西藏植被维护与生态修复、植物多样性重点保护红色名录的制定、特色植物种质资源的保护与利用提供基础数据。
1. 数据与方法
1.1 特有种子植物名录的整理
根据物种分布的地理位置信息,将只分布在西藏自治区境内的种子植物称为西藏特有种子植物。基于专著《中国特有种子植物的多样性及其地理分布》[20]初步确定西藏特有种子植物名录数据,通过查阅Flora of China(http://www.efloras.org/) 、西藏植物志(http://db.kib.ac.cn/XZFLORA/SearchEngine.aspx)、泛喜马拉雅植物志(Flora of Pan-Himalayas)等植物志,结合植物在线数据标本库物种2000中国节点 (http://www.sp2000.org.cn/)、The plant list(TPL) (http://www.theplantlist.org/) 、International Plant Name Index(IPNI)(https://www.ipni.org/) 、Plants of the World online(POWO)(https://powo.science.kew.org/)、Tropicos (https://www.tropicos.org/home)、中国苦苣苔科植物保育中心 (http://gccc.gxib.cn/cn/)等植物数据库网站,再根据近两年中国植物新种文献资料整理结果[21, 22]、检索新物种发表信息[23-26]、专科专属分类修订成果[27]、野外数据资料等的整合,对植物名录进行补充、修正与完善,从而得到较为精准的西藏特有种子植物名录数据库。
1.2 物种多样性组成特征与区系分析
按科属种分类群对西藏特有种子植物物种组成和生活型(乔木、灌木、草本植物等)多样性组成结构进行统计,分析包含物种数最多的科属与生活型构成比例。另外,按照吴征镒等[28]及陈灵芝等[29]对植物区系分布类型的划分原理与方法,确定西藏特有种子植物各属级植物区系分布区类型,并进行区系统计分析。
1.3 多样性分布格局的构建
通过查阅Flora of China ( FOC )( http://www.efloras.org/) 、IPNI、全球生物多样性信息网络(GBIF)(http://www.gbif.org/)、中国数字植物标本馆(CVH)(http://www.cvh.org.cn/)、中国植物图像库(PPBC)(http://ppbc.iplant.cn/)、西藏植物志(http://db.kib.ac.cn/XZFLORA/SearchEngine.aspx)、泛喜马拉雅植物志等植物志及植物在线数据标本库,结合多次野外植物学调查数据和新种文献资料,确定西藏特有种子植物县级分布区及海拔分布范围(假定植物志记载的海拔梯度内物种连续分布)。
按照县级行政区域内特有种子植物分布的物种数量,利用ArcGIS 10.4软件的空间地理制图功能来绘制西藏特有种子植物县域尺度的多样性分布格局图。运用Excel 2019软件对海拔梯度区间内的特有物种数量进行录入和整理,并制作图表。统计每一海拔段内特有物种的数量分布,分析特有物种数量的海拔梯度格局变化趋势,并制作变化趋势图。
2. 结果与分析
2.1 科属种多样性构成
经统计,西藏特有种子植物有1 079种(不包含种下分类等级,见附表1
1 )),占西藏种子植物(8 008种,包含种下等级)[30]的13.5%;占青藏高原特有种子植物(3 764种)[18]的28.7%,表明西藏种子植物特有现象普遍。根据在线网站植物智(http://www.iplant.cn/)和物种2000中国节点 (http://www.sp2000.org.cn/)的分类结果,1 079种西藏特有种子植物隶属于89科295属。其中被子植物1 075种,裸子植物仅有4种,分别占西藏特有种子植物总种数的99.6%和0.4%。裸子植物仅有4种,分别为察隅冷杉(Abies chayuensis Cheng et L. K. Fu)、贡布红杉(Larix kongboensis R. R. Mill)、诚氏麻黄(Ephedra chengiae Y. Yang & D. K. Ferguson)和通麦柏木(Cupressus rushforthii Maerki & J. Hoch)。根据生活型多样性特征,特有种可分为木本植物与草本植物两大类,具体可分为乔木、灌木、木质藤本、多年生草本、一年生草本、附生草本、寄生草本等13种生活型(表1)。包含物种数最多的前7种生活型类型为多年生草本(716种)、灌木(151种)、乔木(65种)、一年生草本(54种)、附生灌木(30种)、附生草本(18种)、亚灌木(16种),分别占西藏特有种子植物总数的66.4%、14.0%、6.0%、5.0%、2.8%、1.7%与1.5%。由此可知,西藏特有种子植物中生活型类型丰富多样,多年生草本植物占优势。木本植物中,灌木类占54.9%,是主要的生活型,其次是乔木,木质藤本类植物最少。草本植物中,多年生草本是主要生活型,占89.1%,二年生草本、攀援草本、全寄生草本与腐生草本所含物种数均小于10,全寄生类植物与腐生草本植物各有1种,分别为大鳞菟丝子(Cuscuta macrolepis R. C. Fang & S. H. Huang)与米林鸟巢兰(Neottia milinensis J. J. Lei & X. H. Jin),生活型丰富多样。
表 1 西藏特有种子植物的生活型Table 1. Growth forms of seed plants endemic to Tibet, China生活型
Growth form物种数
Number of species百分比
Percentages / %木本
植物乔木 65 6.0 灌木 151 14.0 亚灌木 16 1.5 附生灌木 30 2.8 木质藤本 12 1.1 草本
植物多年生草本 716 66.4 二年生草本 7 0.6 一年生草本 54 5.0 攀援草本 7 0.6 附生草本 18 1.7 全寄生草本 1 0.1 腐生草本 1 0.1 从种的数量上看(表2),物种数大于20的有菊科、毛茛科、豆科、报春花科、罂粟科等19个科 ,共包含843种,占西藏特有种子植物总物种数的78.0%,其中菊科、毛茛科、杜鹃花科、豆科、报春花科、禾本科、小檗科所含特有物种数均 ≥ 40种,毛茛科与菊科的物种数更是超过了100个,分布占总物种数的10.0%与11.7%。
表 2 西藏特有含20种以上科种子植物及其所占比例Table 2. Percentages of families containing more than 20 endemic species in Tibet, China科名
Family物种数
Number of species百分比
Percentage / %报春花科 49 4.5 唇形科 25 2.3 豆科 57 5.3 杜鹃花科 65 6.0 禾本科 41 3.8 虎耳草科 37 3.4 菊科 126 11.7 苦苣苔科 24 2.2 兰科 30 2.8 列当科 38 3.5 龙胆科 24 2.2 毛茛科 107 10.0 蔷薇科 31 2.9 伞形科 25 2.3 石竹科 30 2.8 小檗科 42 3.9 杨柳科 22 2.0 罂粟科 48 4.4 紫草科 22 2.0 其他科 236 22.0 包含物种数在10种以上的属及其所占比例的统计结果见表3,分别有翠雀属(Delphinium)、紫堇属(Corydalis)、杜鹃花属(Rhododendron)等24个属,共计530种,占西藏特有种子植物总物种数的48.9%,优势属的现象也很明显。物种数超过30的有小檗属(Berberis)、马先蒿属(Pedicularis)、报春花属(Primula)、杜鹃花属(Rhododendron)、虎耳草属(Saxifraga)、紫堇属(Corydalis)和翠雀属(Delphinium)。此区域也是一些特征属的现代分布与起源演化中心,如马先蒿属,该属中青藏高原特有种为195种,而其中只在西藏分布的有38种(19.5%);虎耳草属西藏特有种为36种,占该属青藏高原特有种(116种)的31.0%;杜鹃花属中青藏高原特有种为159种,其中只分布在西藏的有37种(23.3%);紫堇属包含126个青藏高原特有种,只分布在西藏的就有39种(31.0%)。
表 3 含10种以上属西藏特有种子植物及其所占比例Table 3. Percentages of genera containing more than 10 endemic species in Tibet, China属名
Genus物种数
Number of species百分比
Percentage / %柳属 Salix 20 1.9 楼梯草属Elatostema 13 1.2 蝇子草属Silene 12 1.2 乌头属Aconitum 29 2.8 翠雀属Delphinium 40 3.8 毛茛属Ranunculus 16 1.5 唐松草属Thalictrum 10 1 紫堇属Corydalis 39 3.7 虎耳草属Saxifraga 35 3.4 黄芪属Astragalus 27 2.5 棘豆属Oxytropis 14 1.3 凤仙花属Impatiens 12 1.2 树萝卜属Agapetes 16 1.5 杜鹃花属Rhododendron 37 3.5 报春花属Primula 35 3.4 龙胆属Gentiana 11 1.1 马先蒿属Pedicularis 32 3 小檗属Berberis 40 3.8 蒿属Artemisia 11 1.1 垂头菊属Cremanthodium 12 1.2 橐吾属Ligularia 11 1.1 风毛菊属Saussurea 29 2.8 秋海棠属Begonia 10 1 其他属 532 51 2.2 属的分布区类型结构
对1 079种西藏特有种子植物进行属的分布区类型划分,结果见表4。1 079种植物隶属于297属,其分布区可划分为15个类型和18个变型,包括了吴征镒等[28]确定的中国植物区系的全部15个类型。具体可分为世界分布的属(23属)、热带亚热带分布的属(81属)、温带性质的属(183属)、中国特有分布的属(10属)等4大类,分别占西藏特有种子植物总属的7.7%、27.3%、61.6%与3.4%。
表 4 西藏特有种子植物属的分布类型Table 4. Distribution types of seed plant genera endemic to Tibet, China分布类型 Area type 属数 Number of genera 百分比 Percentage /% 1. 广布 23 7.7 2. 泛热带分布 18 6.1 2-1. 热带亚洲、大洋洲(至新西兰)和中美洲至南美洲(或墨西哥)间断 2 0.7 2-2. 热带亚洲、非洲和中美洲至南美洲间断Disjuncted 2 0.7 3. 热带亚洲和热带美洲间断分布 6 2.0 4. 旧世界热带分布 8 2.7 4-1. 热带亚洲、非洲(或东非、马达加斯加)和大洋洲间断 2 0.7 5. 热带亚洲至热带大洋洲分布 15 5.0 6. 热带亚洲和热带非洲分布 3 1.0 6-2. 热带亚洲和东非或马达加斯加间断 2 0.7 7. 热带亚洲分布 18 6.1 7-1.爪哇(或苏门达腊)、喜马拉雅至华南、西南间断或星散 3 1.0 7-2. 热带印度至华南(特别是滇南) 1 0.3 7-4. 越南(或中南半岛)至华南(或西南) 1 0.3 热带成分(2 ~ 7-4)合计 81 27.3 8. 北温带分布 56 19.0 8-2. 北极-高山 6 2.0 8-4. 北温带和南温带间断分布 22 7.4 8-5. 欧亚和温带南美洲间断 3 1.0 9. 东亚和北美间断分布 10 3.4 9-1. 东亚和墨西哥美洲间断 2 0.7 10. 旧世界温带分布 19 6.4 10-1. 地中海、西亚(或中亚)和东亚间断 1 0.3 10-2. 地中海和喜马拉雅间断 2 0.7 10-3. 欧亚和南部非洲(有时还有大洋洲)间断 1 0.3 11. 温带亚洲分布 6 2.0 12. 地中海、西至中亚分布 1 0.3 13. 中亚分布 4 1.3 13-1. 中亚东部(或中部亚洲)分布 1 0.3 13-2. 中亚至喜马拉雅和华西南 9 3.0 14. 东亚分布 7 2.4 14SH. 中国-喜马拉雅 32 10.8 14SJ. 中国-日本 1 0.3 温带成分(8 ~ 14SJ)合计 183 61.6 15. 中国特有分布 10 3.4 总计 297 100.0 世界分布类型的属中包含毛茛属(Ranunculus)、碎米荠属(Cardamine)、悬钩子属(Rubus)、远志属(Polygala)、鼠尾草属(Salvia)、飞蓬属(Erigeron)、千里光属(Senecio)等大属。
温带性质的属中分布类型及变型丰富多样,尤其以北温带分布类型的属为优势,如柏木属(Cupressus)、冷杉属(Abies)、蔷薇属(Rosa)、百合属(Lilium)与披碱草属(Elymus)等共56属,占西藏特有种子植物总属数的19.0%;其次是中国-喜马拉雅分布变型,包括铁破锣属(Beesia)、丛菔属(Solms-laubachia)、独花报春属(Omphalogramma)、玉山竹属(Yushania)与镰序竹属(Drepanostachyum)等32属,占西藏特有种子植物总属数的10.8%。
热带亚热带性质的属同样丰富多样,以热带亚洲分布类型和泛热带分布类型的属占优,均有18属,占西藏特有种子总属数的6.1%,其次是热带亚洲至热带大洋洲分布的属,为15属,占西藏特有种子总属数的5.0%。泛热带分布类型的有胡椒属(Piper)、花椒属(Zanthoxylum)、榕属(Ficus)、秋海棠属(Begonia)、薯蓣属(Dioscorea)、石豆兰属(Bulbophyllum)与山壳骨属(Pseuderanthemum)等。热带亚洲分布类型有润楠属(Machilus)、坡垒属(Hopea)、线柱苣苔属(Rhynchotechum)、芒毛苣苔属(Aeschynanthus)、树萝卜属(Agapetes)、新小竹属(Neomicrocalamus)、兜兰属(Paphiopedilum)和蛇根草属(Ophiorrhiza)等。热带亚洲至热带大洋洲分布的属有姜属(Zingiber)、蝴蝶兰属(Phalaenopsis)、崖爬藤属(Tetrastigma)、盆距兰属(Gastrochilus)、兰属(Cymbidium)、豆蔻属(Amomum)、小囊兰属(Micropera)、苞舌兰属(Spathoglottis)等。
此外,中国特有分布的属有翅果蓼属(Parapteropyrum)、辐花属(Lomatogoniopsis)、丝苞菊属(Bolocephalus)、秦岭藤属(Biondia)、合头菊属(Syncalathium)、君范菊属(Sinoleontopodium)等,以及2022年才描述发表的单种属十字花科雪古拉荠属(Pulvinatusia)。
2.3 植物多样性分布格局
2.3.1 植物多样性水平地理分布格局
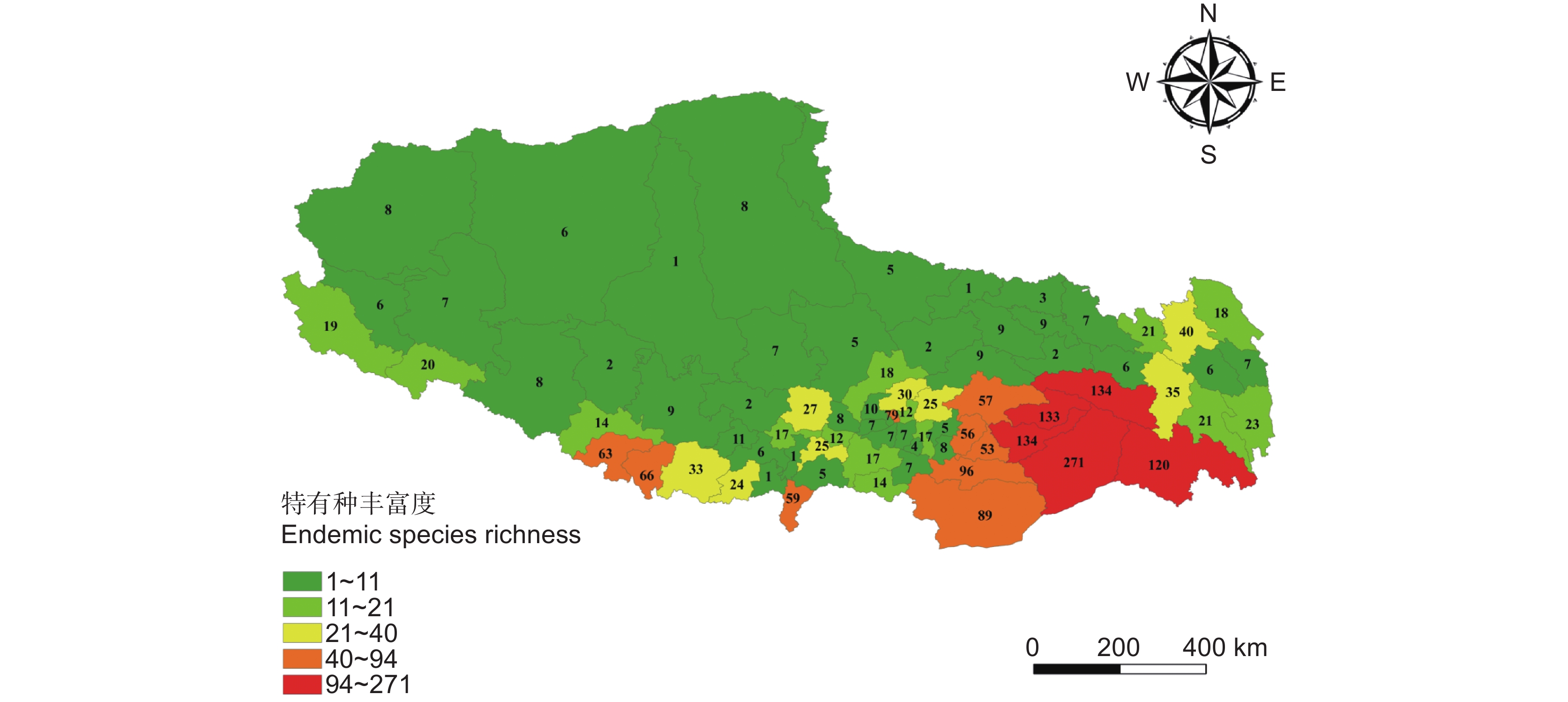
从图1可以看出,西藏特有种子植物总体上呈从东南向西北递减的变化趋势,集中分布在气候温暖湿润的东南部与南部的喜马拉雅几条大沟谷中。物种丰富度呈东南部>南部>中部>东部>西南部>西部>北部的水平多样性地理分布格局,喜马拉雅东南麓是西藏特有种子植物丰富度最高的区域。从县级分布来看,西藏特有种子植物集中分布在东南部的墨脱县、巴宜区、波密县、米林县、察隅县、隆子县、错那县,中部的城关区,以及南部的吉隆县、聂拉木县、亚东县等地区,其中墨脱县特有种数量最多(271种)。特有种数量超过100的有墨脱县、巴宜区、波密县、米林县、察隅县等5个县级行政区。西藏北部和西北部的那曲高原和羌塘高原地区特有种数量较少,物种组成也较为单一,县级分布物种数均不超过10种。西藏地区县级特有种数量之间差异明显,特有种县级分布数量与该区域的植物标本采集强度关系密切。西藏中部的拉萨区域特有种数量明显较高,比周边面积更大、地理环境复杂的县域数量多的多,南部一些县的数量也较少,显然与这些地区的植物标本采集强度及研究程度有着必然的联系。
![]() 图 1 基于县域尺度的西藏特有种子植物多样性分布格局Figure 1. County-level diversity patterns of endemic seed plants in Tibet
图 1 基于县域尺度的西藏特有种子植物多样性分布格局Figure 1. County-level diversity patterns of endemic seed plants in Tibet2.3.2 植物多样性垂直分布格局
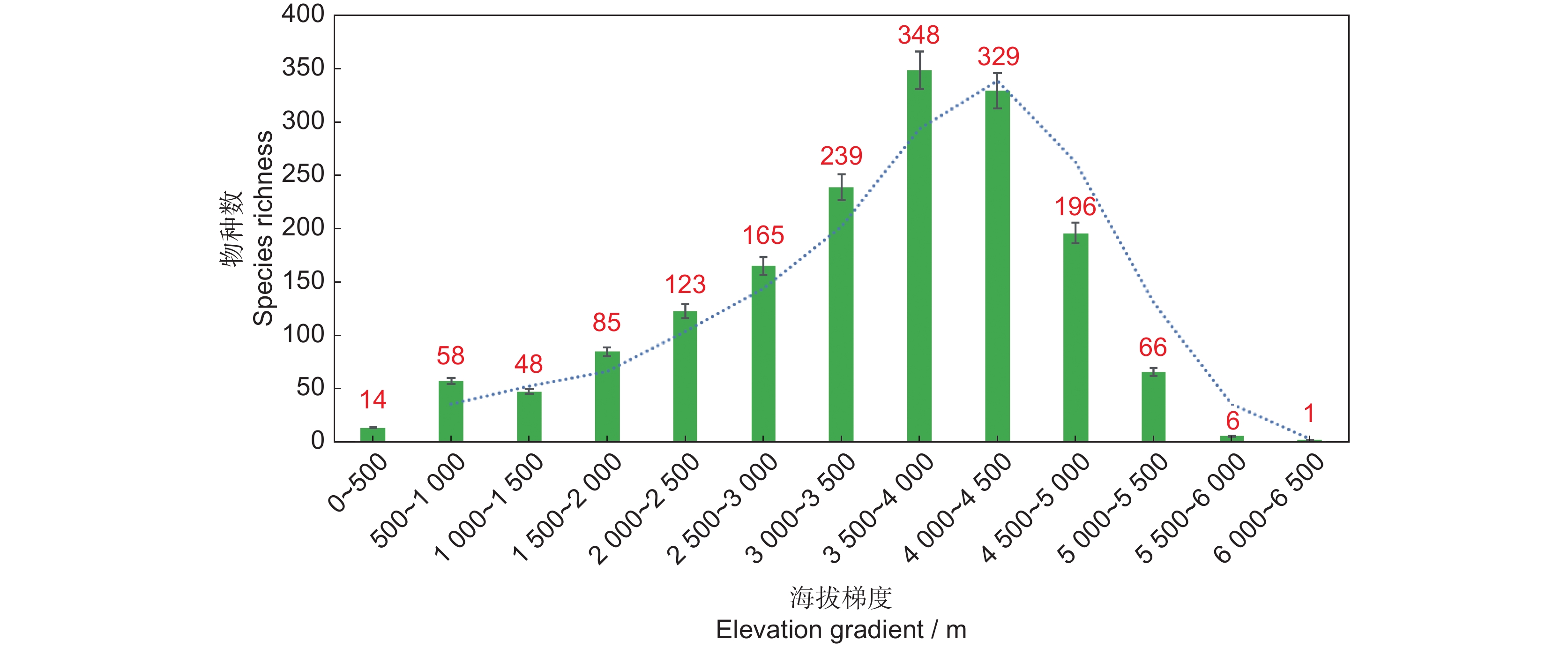
西藏特有种子植物的海拔分布范围极广,跨度从152 m到6400 m。根据海拔梯度,每500 m划分1个区间,从0~500 m至6000~6500 m的物种多样性海拔分布格局如图2所示。结果表明,西藏特有种子植物主要分布在3000~4500 m的中高海拔区间,特有物种丰富度呈单峰分布格局。海拔低于500 m分布的物种只有14种,包括西藏胡椒(Piper arunachalensis Gajurel)、西藏坡垒(Hopea shingkeng (Dunn) Bor)、管花马蓝(Strobilanthes tubiflos (C. B. Clarke) J. R. I. Wood)、粉萼短筒苣苔(Boeica arunachalensis D.Borah, R. Kr. Singh, Taram & A. P. Das)、奥云氏秋海棠(Begonia oyuniae Taram & N. Krishna)等。海拔5 500 m以上分布的有7种,分别是西藏繁缕(Stellaria tibetica Kurz)、仲巴翠雀花(Delphinium chungbaense W. T. Wang)、黑果毛茛(Ranunculus melanogynus W. T. Wang)、金球黄堇(Corydalis chrysosphaera C. Marquand et Airy Shaw)、珠峰鳞蕊芥(Lepidostemon everestianus Al-Shehbaz)、圆叶黄芪(Astragalus orbicularifolius P. C. Li et Ni)、珠峰齿缘草(Eritrichium qofengense Lian et J. Q. Wang)。总体上,随着海拔的升高西藏特有种子植物物种丰富度呈先增加后降低的趋势,即特有物种丰富度的海拔垂直分布格局呈驼峰状模式,符合生物多样性地理分布格局中的法则-中域效应假说[31]。
![]() 图 2 基于海拔梯度的西藏特有种子植物多样性分布格局Figure 2. Elevation gradient-level diversity patterns of endemic seed plants in Tibet
图 2 基于海拔梯度的西藏特有种子植物多样性分布格局Figure 2. Elevation gradient-level diversity patterns of endemic seed plants in Tibet3. 讨论
本研究表明,西藏特有种子植物多样性丰富,依据目前被描述的植物物种,西藏特有种子植物共有1 079种(不包含种下分类等级),占西藏种子植物的13.5%,占青藏高原特有种子植物(3 764种)的28.7%,特有率非常高。物种数最多的前3科为菊科、毛茛科、杜鹃花科;物种数最多的前3属为翠雀属、小檗属、紫堇属。植物生活型多样性分析结果表明,西藏特有种子植物生活型类型丰富多样,有乔木、灌木、木质藤本、多年生草本、一年生草本、附生草本、寄生草本等13种生活型。植物生活型是植物长期对环境适应的表现,是适应各种生态位的结果,这与西藏地区有着丰富的生态环境类型相对应。多年生草本植物占据主要优势,占西藏特有种子植物总物种的66.4%。特有种以多年生草本植物占优势,与其他地区特有种的研究结果相一致[13-16, 32, 33]。西藏特有种子植物的属级地理成分特征为区系类型多样丰富,包含中国种子植物区系的全部15种类型,与各植物区系成分间联系广泛,以温带性质的属为主,其中又以北温带分布性质的属与中国-喜马拉雅成分的属最为密切。另外,在植物区系的发展与起源演化过程中,与热带亚热带性质的属也有紧密的联系和历史渊源,已有研究证明热带性质的属对西藏植物区系的形成与发展有着不容低估的作用[34]。这在一定程度上也反映了西藏植物区系的整体情况。在西藏高原地区独特的地理环境及地质历史事件、一系列山脉隆升等的共同影响下,也孕育出了不少中国特有属的区系成分。
确定物种地理分布格局是生物地理学研究的一个重要出发点[35]。本研究结果表明,西藏特有种子植物的水平地理空间分布(县域尺度)格局极不均匀,特有种沿喜马拉雅山脉集中分布,东南麓是其特有种集中分布的区域,如墨脱县境内分布有271种西藏特有种子植物,其次是巴宜、波密、米林与察隅等县,其特有种丰富度也较高。这些地区独特的新物种很多,且保存有完好的自然生态环境,是探索自然生物资源、生物多样性的宝地。从特有种子植物多样性分布格局研究结果来看,藏东南(墨脱、察隅、波密、米林等地)也是青藏高原特有种丰富度最高的热点区域[18]。西藏特有种子植物的空间地理分布格局与总的种子植物物种丰富度分布格局相一致[36]。从垂直海拔分布格局来看,西藏特有种子植物丰富度随着海拔的升高呈先增后减的变化趋势,特有物种集中分布在3000~4500 m的中高海拔区间。Tiwari 等[37]对尼泊尔喜马拉雅山脉特有植物的分布格局进行了研究,发现这些植物在海拔3800~4200 m之间物种丰富度最高;Werff 和Consiglio[38]对秘鲁分布的特有植物进行了海拔分布格局研究,也发现了类似结果,即特有种的密度在中高海拔出现峰值,与本文的研究结果相一致。在物种海拔垂直分布格局上,相比总的物种丰富度峰值,特有物种的丰富度峰值通常位于更高的海拔区间,这与高海拔地区与世隔绝的加剧形成孤岛效应、土壤表面积的减少、短距离内气候和土壤条件的急剧变化有关,这些因素导致物种种群较小且零散,形成种群隔离,从而有利于狭域特有物种的形成[37, 39, 40]。另外,长期以来,低海拔地区的植物区系和植被受到人类活动的强烈影响,与大范围广布的非特有种相比,特有种可能已受不同程度的影响而灭绝,导致特有物种丰富度在高于总物种丰富度的海拔梯度上出现峰值[5]。
有些特有种的分布范围较广,有些则分布较狭窄,只有1~2个分布点。县域行政区特有种的分布与研究的深入程度及植物标本的采集强度等着有极为密切的关系。西藏中部地区的拉萨比周边几个面积更大、生境类型更复杂的县域高许多,表现出特有种极高的现象,可能与这一原因有关。此地区交通相对便利,自然条件没有那么恶劣,因而是青藏高原植物标本采集活动较为集中的重点地区。这从一定程度上也反映出西藏地区植物标本资源采集的薄弱,甚至还有很多空白地区,但本研究结果在很大程度上也反映了西藏特有种分布格局的整体情况。
本研究基于县域尺度的植物多样性分布格局识别出3个西藏特有种子植物多样性集中丰富的热点地区。Lu等[41]通过重建我国被子植物时间系统发育树,整合物种分布数据、特有类群、濒危性等角度,最后基于综合丰富度指数、综合系统发生指数和受威胁物种分布,识别出42个生物多样性优先保护区,其中西藏有3个,即西藏东南部的雅鲁藏布江-南迦巴瓦峰地区、西藏南部的珠穆朗玛地区与西藏中部的甘巴拉山地区,该结果与本研究相一致。高原面上连续的山脉和峡谷为植物物种迁移与扩散提供了条件,使这些区域的物种丰富度高度集中且多样[42],本研究通过种子植物特有种丰富度识别出的优先保护区域也基本上沿山脉(喜马拉雅山)和峡谷(雅鲁藏布江)分布。
山地热带亚热带潮湿气候的藏东南和藏南地区包含了大量的西藏特有种子植物。未来在资源条件有限的情况下,可针对性地对这些优先区域进行植物多样性调查与保护生物学的研究,同时进一步对每种西藏特有种子植物野外居群的分布范围、种群数量、种群动态特征、生存状况及大尺度地理分布范围、生境异质性等进行全方位实地野外调查,评价特有种的稀有度、濒危等级及保护地位,为制定西藏自治区重点保护植物红色名录的工作提供基础数据,并为国家重点保护野生植物名录的制定提供数据支撑。同时,特有物种是一个地区重要的生物种质资源,建议加大对特有种的实地种群调查和特有珍稀物种的收集保护工作力度,并加强对西藏特有种子植物各个层次的科学研究力度。
1 如需查阅附件内容请登录《植物科学学报》网站(http://www.plantscience.cn)查看本期文章。 -
![]()
图 1 基于县域尺度的西藏特有种子植物多样性分布格局
Figure 1. County-level diversity patterns of endemic seed plants in Tibet
![]()
图 2 基于海拔梯度的西藏特有种子植物多样性分布格局
Figure 2. Elevation gradient-level diversity patterns of endemic seed plants in Tibet
表 1 西藏特有种子植物的生活型
Table 1 Growth forms of seed plants endemic to Tibet, China
生活型
Growth form物种数
Number of species百分比
Percentages / %木本
植物乔木 65 6.0 灌木 151 14.0 亚灌木 16 1.5 附生灌木 30 2.8 木质藤本 12 1.1 草本
植物多年生草本 716 66.4 二年生草本 7 0.6 一年生草本 54 5.0 攀援草本 7 0.6 附生草本 18 1.7 全寄生草本 1 0.1 腐生草本 1 0.1  下载: 导出CSV
下载: 导出CSV
表 2 西藏特有含20种以上科种子植物及其所占比例
Table 2 Percentages of families containing more than 20 endemic species in Tibet, China
科名
Family物种数
Number of species百分比
Percentage / %报春花科 49 4.5 唇形科 25 2.3 豆科 57 5.3 杜鹃花科 65 6.0 禾本科 41 3.8 虎耳草科 37 3.4 菊科 126 11.7 苦苣苔科 24 2.2 兰科 30 2.8 列当科 38 3.5 龙胆科 24 2.2 毛茛科 107 10.0 蔷薇科 31 2.9 伞形科 25 2.3 石竹科 30 2.8 小檗科 42 3.9 杨柳科 22 2.0 罂粟科 48 4.4 紫草科 22 2.0 其他科 236 22.0
下载: 导出CSV
表 3 含10种以上属西藏特有种子植物及其所占比例
Table 3 Percentages of genera containing more than 10 endemic species in Tibet, China
属名
Genus物种数
Number of species百分比
Percentage / %柳属 Salix 20 1.9 楼梯草属Elatostema 13 1.2 蝇子草属Silene 12 1.2 乌头属Aconitum 29 2.8 翠雀属Delphinium 40 3.8 毛茛属Ranunculus 16 1.5 唐松草属Thalictrum 10 1 紫堇属Corydalis 39 3.7 虎耳草属Saxifraga 35 3.4 黄芪属Astragalus 27 2.5 棘豆属Oxytropis 14 1.3 凤仙花属Impatiens 12 1.2 树萝卜属Agapetes 16 1.5 杜鹃花属Rhododendron 37 3.5 报春花属Primula 35 3.4 龙胆属Gentiana 11 1.1 马先蒿属Pedicularis 32 3 小檗属Berberis 40 3.8 蒿属Artemisia 11 1.1 垂头菊属Cremanthodium 12 1.2 橐吾属Ligularia 11 1.1 风毛菊属Saussurea 29 2.8 秋海棠属Begonia 10 1 其他属 532 51
下载: 导出CSV
表 4 西藏特有种子植物属的分布类型
Table 4 Distribution types of seed plant genera endemic to Tibet, China
分布类型 Area type 属数 Number of genera 百分比 Percentage /% 1. 广布 23 7.7 2. 泛热带分布 18 6.1 2-1. 热带亚洲、大洋洲(至新西兰)和中美洲至南美洲(或墨西哥)间断 2 0.7 2-2. 热带亚洲、非洲和中美洲至南美洲间断Disjuncted 2 0.7 3. 热带亚洲和热带美洲间断分布 6 2.0 4. 旧世界热带分布 8 2.7 4-1. 热带亚洲、非洲(或东非、马达加斯加)和大洋洲间断 2 0.7 5. 热带亚洲至热带大洋洲分布 15 5.0 6. 热带亚洲和热带非洲分布 3 1.0 6-2. 热带亚洲和东非或马达加斯加间断 2 0.7 7. 热带亚洲分布 18 6.1 7-1.爪哇(或苏门达腊)、喜马拉雅至华南、西南间断或星散 3 1.0 7-2. 热带印度至华南(特别是滇南) 1 0.3 7-4. 越南(或中南半岛)至华南(或西南) 1 0.3 热带成分(2 ~ 7-4)合计 81 27.3 8. 北温带分布 56 19.0 8-2. 北极-高山 6 2.0 8-4. 北温带和南温带间断分布 22 7.4 8-5. 欧亚和温带南美洲间断 3 1.0 9. 东亚和北美间断分布 10 3.4 9-1. 东亚和墨西哥美洲间断 2 0.7 10. 旧世界温带分布 19 6.4 10-1. 地中海、西亚(或中亚)和东亚间断 1 0.3 10-2. 地中海和喜马拉雅间断 2 0.7 10-3. 欧亚和南部非洲(有时还有大洋洲)间断 1 0.3 11. 温带亚洲分布 6 2.0 12. 地中海、西至中亚分布 1 0.3 13. 中亚分布 4 1.3 13-1. 中亚东部(或中部亚洲)分布 1 0.3 13-2. 中亚至喜马拉雅和华西南 9 3.0 14. 东亚分布 7 2.4 14SH. 中国-喜马拉雅 32 10.8 14SJ. 中国-日本 1 0.3 温带成分(8 ~ 14SJ)合计 183 61.6 15. 中国特有分布 10 3.4 总计 297 100.0
下载: 导出CSV
-
[1] 黄继红,张金龙,杨永,马克平. 特有植物多样性分布格局测度方法的新进展[J]. 生物多样性,2013,21(1):99−110. doi: 10.3724/SP.J.1003.2013.12175 Huang JH,Zhang JL,Yang Y,Ma KP. Advances in methods for measuring patterns of endemic plant diversity[J]. Biodiversity Science,2013,21 (1):99−110. doi: 10.3724/SP.J.1003.2013.12175
[2] Stoch F. How many endemic species? Species richness assessment and conservation priorities in Italy[J]. Belg J Entomol,2000,2 (1):125−133.
[3] Cowling RM,Samways MJ. Predicting global patterns of endemic plant species richness[J]. Biodiversity Lett,1994,2 (5):127−131. doi: 10.2307/2999776
[4] Burlakova LE,Karatayev AY,Karatayev VA,May ME,Bennett DL,Cook MJ. Endemic species:contribution to community uniqueness,effect of habitat alteration,and conservation priorities[J]. Biol Conserv,2011,144 (1):155−165. doi: 10.1016/j.biocon.2010.08.010
[5] Trigas P,Tsiftsis S,Tsiripidis I,Iatrou G. Distribution patterns and conservation perspectives of the endemic flora of Peloponnese (Greece)[J]. Folia Geobot,2012,47 (4):421−439. doi: 10.1007/s12224-012-9130-4
[6] Clements R,Ng PKL,Lu XX,Ambu S,Schilthuizen M,Bradshaw CJA. Using biogeographical patterns of endemic land snails to improve conservation planning for limestone karsts[J]. Biol Conserv,2008,141 (11):2751−2764. doi: 10.1016/j.biocon.2008.08.011
[7] Essl F,Staudinger M,Stöhr O,Schratt-Ehrendorfer L,Rabitsch W,Niklfeld H. Distribution patterns,range size and niche breadth of Austrian endemic plants[J]. Biol Conserv,2009,142 (11):2547−2558. doi: 10.1016/j.biocon.2009.05.027
[8] Georghiou K,Delipetrou P. Patterns and traits of the endemic plants of Greece[J]. Bot J Linn Soc,2010,162 (2):130−153. doi: 10.1111/j.1095-8339.2010.01025.x
[9] Myers N,Mittermeier RA,Mittermeier CG,da Fonseca GAB,Kent J. Biodiversity hotspots for conservation priorities[J]. Nature,2000,403 (6772):853−858. doi: 10.1038/35002501
[10] Schmeller DS,Gruber B,Budrys E,Framsted E,Lengyel S,Henle K. National responsibilities in European species conservation:a methodological review[J]. Conserv Biol,2008,22 (3):593−601. doi: 10.1111/j.1523-1739.2008.00961.x
[11] Darwall WRT,Vié JC. Identifying important sites for conservation of freshwater biodiversity:extending the species‐based approach[J]. Fish Manage Ecol,2005,12 (5):287−293. doi: 10.1111/j.1365-2400.2005.00449.x
[12] Knight AT,Cowling RM,Rouget M,Balmford A,Lombard AT,Campbell BM. Knowing but not doing:selecting priority conservation areas and the research–implementation gap[J]. Conserv Biol,2008,22 (3):610−617. doi: 10.1111/j.1523-1739.2008.00914.x
[13] 季春峰,钱萍,杨清培,杨光耀,裘利洪,等. 江西特有植物区系、地理分布及生活型研究[J]. 武汉植物学研究,2010,28(2):153−160. Ji CF,Qian P,Yang QP,Yang GY,Qiu LH,et al. Research on the floristic characteristics,geographical distributions and life-forms of plants endemic to Jiangxi Province[J]. Journal of Wuhan Botanical Research,2010,28 (2):153−160.
[14] 丁莉,唐文秀,骆文华,盘波,韦毅刚,黄仕训. 广西特有植物区系特征研究[J]. 广西植物,2010,30(2):202−208. Ding L,Tang WX,Luo WH,Pan B,Wei YG,Huang SX. Floristic characteristics of endemic plants to Guangxi[J]. Guihaia,2010,30 (2):202−208.
[15] 曹晓杰,曹伟,张悦,郭佳,高燕. 中国东北地区植物特有性研究[J]. 植物研究,2022,42(5):753−761. Cao XJ,Cao W,Zhang Y,Guo J,Gao Y. Plant endemism in northeast China[J]. Bulletin of Botanical Research,2022,42 (5):753−761.
[16] 董洪进,周亚东,王青锋. 湖北特有种子植物分布特征分析[J]. 植物科学学报,2016,34(5):705−714. Dong HJ,Zhou YD,Wang QF. Analysis on the distribution patterns of seed plants endemic to Hubei[J]. Plant Science Journal,2016,34 (5):705−714.
[17] 朱青,盛岩. 滇西北丽江市中国种子植物特有种组成分析[J]. 浙江农林大学学报,2019,36(3):479−485. Zhu Q,Sheng Y. Composition of seed plants endemic to China in Lijiang City,Northwest Yunnan[J]. Journal of Zhejiang A& F University,2019,36 (3):479−485.
[18] 于海彬,张镱锂,刘林山,陈朝,祁威. 青藏高原特有种子植物区系特征及多样性分布格局[J]. 生物多样性,2018,26(2):130−137. doi: 10.17520/biods.2017078 Yu HB,Zhang YL,Liu LS,Chen C,Qi W. Floristic characteristics and diversity patterns of seed plants endemic to the Tibetan Plateau[J]. Biodiversity Science,2018,26 (2):130−137. doi: 10.17520/biods.2017078
[19] 吴征镒. 西藏植物志(第5卷)[M]. 北京: 科学出版社, 1987: 874-877. [20] 黄继红, 马克平, 陈彬. 中国特有种子植物的多样性及其地理分布[M]. 北京: 高等教育出版社, 2014: 22. [21] 杜诚,刘军,叶文,廖帅,葛斌杰,等. 中国植物新分类群、新名称 2020 年度报告[J]. 生物多样性,2021,29(8):1011−1020. doi: 10.17520/biods.2021122 Du C,Liu J,Ye W,Liao S,Ge BJ,et al. Annual report of new taxa and new names for Chinese plants in 2020[J]. Biodiversity Science,2021,29 (8):1011−1020. doi: 10.17520/biods.2021122
[22] 杜诚,刘军,叶文,廖帅. 中国植物新分类群、新名称变化2021年年度报告[J]. 生物多样性,2022,30(8):22207. doi: 10.17520/biods.2022207 Du C,Liu J,Ye W,Liao S. 2021 annual report on new taxa and nomenclatural changes of Chinese plants[J]. Biodiversity Science,2022,30 (8):22207. doi: 10.17520/biods.2022207
[23] Meng SY,Wang ZM,Ye L. Integrative taxonomy supports two new species of Rhodiola (Crassulaceae) in Xizang,China[J]. Diversity,2022,14 (4):289. doi: 10.3390/d14040289
[24] Yang B,Ya JD,Tong YH,Wang PY,Liu C,et al. Agapetes huangiana (Ericaceae),a new species from Southeast Xizang,China[J]. Taiwania,2022,67 (2):254−259.
[25] Li M,Tian CF,Song YF,Ju WB,Pathak ML,Xu B. Stellaria motuoensis (Caryophyllaceae),a new species from Xizang,China[J]. Nord J Bot,2022,2022 (9):e03683. doi: 10.1111/njb.03683
[26] Tong YH,Sun M,Wang BM,Tian HZ. Goodyera medogensis (Orchidaceae),a new species from Tibet,China[J]. PhytoKeys,2022,189:141−154. doi: 10.3897/phytokeys.189.77374
[27] 王文采. 中国银莲花属新分类[J]. 广西植物,2021,41(S1):1−118. Wang WC. A new classification of Anemone (Ranunculaceae) of China[J]. Guihaia,2021,41 (S1):1−118.
[28] 吴征镒, 周浙昆, 孙航, 李德铢, 彭华. 种子植物分布区类型及其起源和分化[M]. 昆明: 云南科技出版社, 2006: 1-532. [29] 陈灵芝, 孙航, 郭柯. 中国植物区系与植被地理[M]. 北京: 科学出版社, 2015: 30-163. [30] The Biodiversity Committee of Chinese Academy of Sciences. Catalogue of Life China: 2022 Annual Checklist[DB/OL]. Beijing, China, 2022.
[31] Colwell RK,Lees DC. The mid-domain effect:geometric constraints on the geography of species richness[J]. Trends Ecol Evol,2000,15 (2):70−76. doi: 10.1016/S0169-5347(99)01767-X
[32] 孙冠花. 祁连山国家公园(青海片区)中国特有植物多样性及其保护利用[D]. 杨凌: 西北农林科技大学, 2021: 15-16. [33] 丁博,唐粒,刘冬梅,王茜,邓洪平. 重庆特有种子植物区系特征研究[J]. 西北植物学报,2016,36(4):825−830. Ding B,Tang L,Liu DM,Wang Q,Deng HP. Floristic characteristics of endemic seed plants in Chongqing[J]. Acta Bot Boreal Occident Sin,2016,36 (4):825−830.
[34] 李恒,武素功. 西藏植物区系区划和喜马拉雅南部植物地区的区系特征[J]. 地理学报,1983,38(3):252−261. Li H,Wu SG. The regionalization of Xizang (Tibet) flora and the floristic structure of south Himalaya region[J]. Acta Geographica Sinica,1983,38 (3):252−261.
[35] Linder HP. On areas of endemism,with an example from the African Restionaceae[J]. Syst Biol,2001,50 (6):892−912. doi: 10.1080/106351501753462867
[36] 杨光澍,冯建孟. 西藏地区种子植物多样性分布格局和多样性分化特点[J]. 信阳师范学院学报(自然科学版),2016,29(3):405−411. Yang GS,Feng JM. The spatial patterns of species diversity of seed plants and its differentiation in Tibet[J]. Journal of Xinyang Normal University (Natural Science Edition)
,2016,29 (3):405−411. [37] Tiwari A,Uprety Y,Rana SK. Plant endemism in the Nepal Himalayas and phytogeographical implications[J]. Plant Diversity,2019,41 (3):174−182. doi: 10.1016/j.pld.2019.04.004
[38] Van der Werff H,Consiglio T. Distribution and conservation significance of endemic species of flowering plants in Peru[J]. Biodiversity Conserv,2004,13 (9):1699−1713. doi: 10.1023/B:BIOC.0000029334.69717.f0
[39] Ríos-Touma B,Cuesta F,Rázuri-Gonzales E,Holzenthal R,Tapia A,Calderón-Loor M. Elevational biodiversity gradients in the Neotropics:perspectives from freshwater caddisflies (Insecta:Trichoptera)[J]. PLoS One,2022,17 (8):e0272229. doi: 10.1371/journal.pone.0272229
[40] Kessler M. The elevational gradient of Andean plant endemism:varying influences of taxon‐specific traits and topography at different taxonomic levels[J]. J Biogeogr,2002,29 (9):1159−1165. doi: 10.1046/j.1365-2699.2002.00773.x
[41] Lu LM, Zhao LN, Hu HH, Liu B, Yang YC, et al. A comprehensive evaluation of flowering plant diversity and conservation priority for national park planning in China[J]. Fundam Res, 2022, doi: 10.1016/j.fmre.2022.08.008.
[42] Yu HB,Zhang YL,Wang ZF,Liu LS,Chen Z,Wi W. Diverse range dynamics and dispersal routes of plants on the Tibetan Plateau during the late Quaternary[J]. PLoS One,2017,12 (5):e0177101. doi: 10.1371/journal.pone.0177101
-
期刊类型引用(2)
1. 贺文琪,柴国柱,薛矗,山晓丹,兰小中. 西藏11种菊科药用植物染色体核型研究. 种子. 2025(02): 190-201 .  百度学术
百度学术
2. 马启奥,孙楷,王宏超,李利平,李颖,李晓琳,程蒙,杨光,池秀莲. 高黎贡山区域中药材药性的组成特征及垂直格局. 中国现代中药. 2024(11): 1833-1842 . 百度学术
其他类型引用(5)
-
其他相关附件
-
PDF格式
2023年 5期 王俊伟 附表 点击下载(477KB)
-
计量
- 文章访问数: 293
- HTML全文浏览量: 53
- PDF下载量: 67
- 被引次数: 7
