
Research progress on the involvement of ABCG transporters in the regulation of plant male fertility
-
摘要:
利用雄性不育系制种是利用杂种优势的最有效的途径,尤其对小麦(Triticum aestivum L.)、水稻(Oryza sativa L.)等自花授粉作物的杂种优势利用具有十分重要的意义。ABCG(ATP-binding cassette G transporter)转运蛋白属于ABC(ATP-binding cassette transporter)转运蛋白中的一个大亚族,参与植物各种生物过程,包括调控雄性育性。本文概述了ABC转运蛋白的特点与分类,并系统阐述了ABCG转运蛋白参与调控植物花粉壁和花药角质层发育的研究现状,同时也对ABCG转运蛋白目前研究所存在的问题提出了建议,并对后续的研究进行了展望。
Abstract:The utilization of male sterile lines for seed production represents an effective strategy for exploiting heterosis in self-pollinating crops such as wheat and rice. ABCG transporters, a large subfamily of ABC transporters, play integral roles in various biological processes, including the regulation of male fertility. This review examines the structural characteristics of ABC transporters, emphasizes the critical role of ABCG transporters in regulating the development of the pollen wall and anther cuticle, and discusses the current state of research on ABCG transporters, along with future research prospects. The aim of this review is to enhance our understanding of the involvement of ABCG transporters in the regulation of plant male sterility, thereby providing a theoretical basis for the cross-breeding of self-pollinated crops.
-
Keywords:
- ABCG transporters /
- Male fertility /
- Anther cuticle /
- Pollen wall
-
森林空间结构是根据林木的空间位置和属性,来描述林木间相互关系的空间排列方式。而林木个体的空间分布状态,即分布格局,是研究林木在水平空间相互关系的重要指标,同时也是空间结构的基本特征之一[1]。分布格局的研究方法主要有样方法和距离法,但两种方法均存在局限性。
1999年,有学者提出了由参照树和相邻木组成的描述林分空间结构单元的方法[2]。该方法把林区内任意一棵树和距其最近的4棵相邻树组成的结构小组称为林分空间结构单元。角尺度(Wi)就是基于林分空间结构单元所构建的一个参数,用于描述林木的分布格局[3]。角尺度法主要是通过判断由参照树和相邻木构成的夹角大小,来判断相邻木围绕参照树的均匀性。该方法既可以通过图形来判断分布格局,也可以通过数值来判断,其操作方便,计算简单[4]。林木分布格局在林木和森林的生长演替过程中均发挥直接作用。因此,通过分析森林林分整体的角尺度分布特征,能有效地确定林分演替阶段,了解林分的稳定性。同时,对优势种分布格局进行分析,还能了解优势种与其他物种的关系,如竞争、共生、互助等。
随机木是指Wi=0.50的林木,随机木对应的结构单元称为随机体。该概念是Zhang和Hui[5]在2021年基于角尺度参数提出的。随机木在反映林分整体树种组成和竞争情况等方面具有重要作用。研究发现,随机体比例在天然林中不受地域分布、树种组成、林分结构、格局类型和树木竞争的影响,并由此提出了“随机体-稳定性”假说[6]。但在喀斯特区域,因其特有的“富钙、干旱、瘠薄”等异质生境,加之喀斯特复杂的地质地貌和环境因素,使其成为环境异质性与生物物种多样性的独特响应区。因此,生境异质性强的喀斯特地区天然林中,“随机体-稳定性”假说是否仍然成立,还有待检验。Li等[7]分析了喀斯特地区土壤和岩石两种立地上,不同径阶树木群体以及每种立地上两种生活型树木的随机体分布特征,结果发现随机体的比例和分布受生活型和生境异质性的影响,而与树的大小无关。然而,先前对于随机体在喀斯特地区影响因素的分析,均是以尺度较小的样地作为研究对象,在更大的森林样地尺度中,生活型是否对随机体有影响还不清楚。生境异质性的产生主要有两个原因,一是组成生境的能量和物质在时间或空间上的差异,如生物因素、水分状况等[8],二是区域生态系统运动发展的不平衡性[9]。而不同的环境因子在喀斯特地区中对随机体的影响还未知。此外,立地条件对于林木生长也是一个重要因素[10]。因此,研究喀斯特生境中不同因子对随机体的影响差异具有重要意义。
我国桂西南地区是北热带喀斯特季节性雨林的重要分布区之一,不但包含了从山顶到山坡到洼地的各种地形要素,还包含了水平方向上的高异质性以及垂直方向上的多层性,形成了一系列典型的“峰丛-洼地”生境类型[11, 12]。在喀斯特岩溶山地的地形地貌中,存在着复杂多样的小生境,这些小生境受不同因素的影响而呈现出多样性。此外,在不同的地段、坡向和坡位上,小生境的资源分配和植物组成等方面均存在明显差异。本文基于弄岗喀斯特季节性雨林15-hm2样地的第2次复查数据,从林分整体和优势种角尺度分布特征,以及生活型、生物因子和地形因子对随机体的影响进行分析。研究旨在探讨:(1)基于北热带喀斯特季节性雨林相邻木林分整体和优势种的随机体分布特征;(2)不同生活型树种在空间尺度较大的喀斯特生境中对随机体是否有影响;(3)北热带喀斯特季节性雨林不同环境因子对随机体的影响差异。
1. 材料与方法
1.1 研究区概况
弄岗国家级自然保护区地处广西龙州县与宁明县的交界地带,分弄岗、隆呼、陇瑞3个区,总面积为10 077 hm2。保护区土壤类型为原始石灰土、棕色石灰土、黑色石灰土等。年降水集中在5-9月,降水量分布不均匀,干湿交替较强;年平均温度22 ℃,最低温度13 ℃,最高温度37 ℃~39 ℃。
1.2 样地设置与调查
北热带喀斯特季节性雨林地处弄岗自然保护区弄岗片的弄姆皇(22°25′N,106°57′E)。整个样地南北宽300 m,东西长500 m,海拔为180~370 m,平均海拔260 m,样地坡度范围在3.7°~78.9°,平均坡度41.7°。整个样地生境多样化,属于喀斯特“峰丛-洼地”典型的生境类型,在中国森林生物多样性监测网络中,该样地是唯一的典型热带喀斯特森林监测样地。
根据CTFS全球森林生物多样性监测标准,利用全站仪将15-hm2样地划分成1 500个10 m×10 m的样方。在此基础上,于2011年完成了首次植被调查,对样方内胸径(DBH)≥1 cm个体的名称、胸径、坐标及生长状态等进行了调查并记录。之后,每5年进行一次调查,并于2021年完成第2次复查。本文以2021年的复查数据为依据,选取DBH≥5 cm的个体作为研究对象。
1.3 树种优势度的计算
树种优势度(Dsp)指树种在林分中的数量优势度和空间上的优势度,选取优势度位于前10的物种作为优势种,Dsp的计算公式为:
Dsp=√Dg⋅(1−¯Usp) (1) 式中,Dg表示相对显著度,
ˉUsp 表示树种大小比数均值。Dsp接近1表示非常优势,接近0表示几乎没有优势[13]。树种大小比数均值表达式如下:
¯Usp=1Nsp∑Nspi=1Ui (2) 式中,Nsp表示所观察树种的参照树数量,Ui为树种的第i株树的大小比数值。
1.4 角尺度计算
角尺度(Wi)指的是参照树与4株最近邻木间小于标准角的夹角占比,用下式表示[3]:
Wi=1n∑nj=1Zij (3) 其中:Zij=
1{1(当j个角小于标准角α0)0(否则) 式中,Wi为参照树i的角尺度,n指相邻木个体数,n=4。Zij为参照树i与其相邻木j之间的第j个夹角α与标准角α0之间的大小关系,有两种可能的值,当Zij=1时,表明第j个α角小于标准角,当Zij=0时,表明第j个α角大于标准角。
角尺度均值(
¯W )能够反映一个林分整体的分布情况。其计算公式如下:¯W=1N∑Niwi (4) 优势种角尺度均值用下式表示:
¯Wsp=1Nsp∑Nspi=1 (5) 式中,N为样地林木总株数。当角尺度均值在[0.457,0.517]时为随机分布;大于0.517为聚集分布;小于0.475为均匀分布[14]。角尺度等于0.50的林木被称为随机木,相应林木的结构单元称为随机体[15]。角尺度为0.75或1的林木为聚集木,相应林木结构单元称为聚集体。角尺度为0或0.25的林木被称作均匀木,相应林木的结构单元被称作均匀体。本研究在计算林分整体和优势种角尺度时,设置了5 m缓冲区,并将缓冲区内的林木做相邻木处理,以此消除林木的边缘效应造成的影响。
1.5 Pearson 相关分析
运用Pearson相关分析法,判断随机体与环境因子及环境因子间的相关性。相关系数r计算公式如下:
r=∑(x−ˉx)−(y−ˉy)∑(x−ˉx)2∑(y−ˉy)2 (6) r取值范围为−1~1,当|r|≥0.8,表示两者高度相关;0.5≤|r|<0.8,为中度相关;0.3≤|r|<0.5,表示低相关;当|r|<0.3,表示相关性非常微弱,可视为没有相关性,P<0.05表明有统计学意义。
1.6 环境因子获取
用全站仪获取精确海拔数据,计算每个20 m×20 m样方的平均海拔、坡向、坡度和凹凸度4个地形指标[16-18]。土壤湿度状况以地形湿润指数和干旱度指数为表征,参见Punchi-Manage等[19]的研究。样地内岩石裸露情况采用踏查法进行评估。综上共有7个地形因子。此外,还考查了每个样方内最大胸径、胸高断面积之和、植株个体数和平均胸径4个生物因子,共11个环境因子。
1.7 冗余分析
冗余分析的响应变量由每个样方的随机体个数组成。为了消除林木边缘效应的影响,设置2 m缓冲区,缓冲区内的林木做相邻木处理。解释变量数据包括11个环境因子。冗余分析前,对数据进行标准化处理,并比较自变量的相关系数是否大于7,以此衡量自变量间的共线性。再计算每个变量的方差膨胀因子,当方差膨胀因子<10时,表示无明显的共线性问题[20],可进行冗余分析。采用层次分割方法对冗余分析结果进行变差分解,以获取每个解释变量的校正解释率。
1.8 数据处理
数据分析计算均利用R 4.3.1软件完成,冗余分析在vegan程序包中进行,层次分割采用rdacca.hp程序包[21],利用ForestSAS程序包计算角尺度和大小比数均值[22],Pearson相关系数图在corrplot包中绘制,角尺度分布特征图在ggplot2包中完成,单独效应比例图在Origin 2021软件中绘制。
2. 结果与分析
2.1 林分整体和优势种随机体的比例和分布特征
树种优势度位于前10的物种在森林中起关键作用,尤其是闭花木(Cleistanthus sumatranus (Miq.) Muell. Arg.)、苹婆(Sterculia monosperma Ventenat)和广西牡荆(Vitex kwangsiensis P'ei),不仅在数量上占林分整体的绝大多数,其优势度也是最高的,分别为0.262、0.260、0.245,其他树种的优势度均小于0.200。角尺度计算结果表明,林分整体和优势种角尺度均值都在随机分布范围内(表1)。但林分整体和优势种随机木的分布频率均超过0.5,而均匀木和聚集木分布频率均比随机木低,说明样地中大多数林木的最近相邻木处于随机分布,各个优势种的最近相邻木处于随机分布的也占大多数,即随机体占绝对分布(图1)。
表 1 北热带喀斯特季节性雨林树种优势度排名前10的树种Table 1. Top 10 dominant species in northern tropical karst seasonal rainforest树种
Species科名
Family株数
Number of
plants大小比数
Dominance相对显著度
Relative
significance优势度
Advantage
degree角尺度
Uniform angle
index广西牡荆
Vitex kwangsiensis P'ei唇形科 1 645 0.325 0.122 0.286 0.504 苹婆
Sterculia monosperma Ventenat梧桐科 2 426 0.491 0.134 0.262 0.498 闭花木
Cleistanthus sumatranus (Miq.) Muell. Arg.大戟科 3 054 0.530 0.139 0.256 0.504 劲直刺桐
Erythrina stricta Roxb.豆科 242 0.074 0.031 0.171 0.485 蚬木
Excentrodendron tonkinense (A. Chev.)
H. T. Chang et R. H. Miau锦葵科 483 0.367 0.039 0.156 0.498 海南椴
Diplodiscus trichosperma (Merrill) Y. Tang锦葵科 456 0.317 0.033 0.150 0.509 黄梨木
Boniodendron minus (Hemsl.) T. Chen无患子科 404 0.314 0.026 0.133 0.506 对叶榕
Ficus hispida L. f.桑科 676 0.602 0.031 0.110 0.497 假玉桂
Celtis timorensis Span大麻科 414 0.509 0.020 0.099 0.495 鱼骨木
Psydrax dicocca Gaertn.茜草科 252 0.405 0.015 0.094 0.510 林分整体 − 10 052 0.489 − − 0.500 ![]() 图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.
图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.2.2 不同生活型树种随机体的比例和分布特征
乔木角尺度均值在随机分布范围内,为0.499,随机体占比为57.167%,均匀体占比较小,为23.189%,聚集体次之,为19.644%。灌木角尺度均值为0.519,为聚集分布,随机体占比为59.173%,均匀体占比为19.064%,聚集体为21.763%。乔木和灌木的随机体比例均超过55%,但灌木的聚集体占比高于均匀体。而在乔木中,均匀体占比高于聚集体,表明不同生活型的植物,其随机体比例和分布存在一定的差异(表2)。
表 2 北热带喀斯特季节性雨林不同生活型随机体比例和分布Table 2. Proportion and distribution of random framework among different life forms in northern tropical karst seasonal rainforest生活型
Life style角尺度 Uniform angle index / % 非常均匀
Very regular均匀
Regular随机
Random聚集
Clustered非常聚集
Very clustered角尺度均值
Uniform angle index mean乔木 0.773 22.416 57.167 15.785 3.859 0.499 灌木 0.719 18.345 59.173 16.187 5.576 0.519 2.3 Pearson相关性分析
研究结果显示,随机体与胸高断面积之和、平均海拔、坡度、岩石裸露率、凹凸度、干旱度指数、原有植株个体数之间均存在显著正相关,其中相关性最高的为原有植株个体数,相关系数达0.86;平均海拔和坡度分别与随机体存在低度相关,相关系数分别为0.36和0.31。随机体分别与最大胸径、平均胸径和地形湿润指数存在显著负相关,但与坡向无显著相关性。胸高断面积和最大胸径、坡度和地形湿润指数间也具有较高的相关性,相关系数分别为0.7和−0.72(图2)。
![]() 图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.
图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.2.4 环境因子与随机体的冗余分析及单个解释变量的相对重要性
结合环境因子的相关性和方差膨胀因子分析,将胸高断面积之和剔除。对随机体与生物因子和地形因子进行冗余分析,10个环境因子对随机体的解释量R2经校正后为0.729。其中,生物因子解释了65.6%,地形因子解释了7.3%。未被解释的部分占27.1%。根据层次分割法各环境因子单独效应比例排序,随机体的主要影响因素包括原有植株个体数、平均胸径、平均海拔、地形湿润指数和坡度;贡献较小的是最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数(图3)。
![]() 图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest
图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest3. 讨论
3.1 随机体的分布特征
林木的空间分布格局主要分为随机分布、规则分布和聚集分布,最理想的是随机分布,而随机分布也是天然原始顶极群落中最为显著的结构特征[23]。本研究中,林分整体为随机分布,优势种的角尺度均值也在随机分布范围内,且随机体占比均超过50%。因此,随机体以相似的比例出现在不同树种中,且支持了“随机体-稳定性”假说,与前人研究结果一致[5]。这种结构体相对来说具有较小的生存压力,稳定性高,在竞争情况下不易出现弱势或者不健康的林木,在自然演替过程中存活几率更大[6]。而均匀体和聚集体占比较小,与袁星明等[24]的报道相似。
不同空间分布格局分析方法可能导致结果上的差异。前人研究采用完全随机零模型和泊松异质性零模型对该地区分布格局进行了分析,完全随机零模型结果表明,在小尺度上所有物种均呈聚集分布[25],与本研究结果不一致;然而,当泊松异质性零模型去除生境异质性后,大部分物种表现为随机分布,与本文结果一致。完全随机零模型是基于假设个体随机分布,不受任何因素的干扰而对分布格局进行拟合分析。但在生境异质性强的北热带喀斯特地区,物种的分布显然会受到各种因素的影响。因此,在不考虑任何因素干扰情况下大多数物种呈聚集分布,与本研究结果存在差异。泊松异质性零模型主要用于模拟生境关联,本研究基于相邻木的角尺度法分析林木分布格局,也未直接考虑生境异质性,所以二者结果一致。另外,角尺度法不存在尺度效应,而空间点格局法可以拟合分析不同尺度的林木分布格局,二者各具特点。对于这两种方法,有学者也进行了对比,发现角尺度法在判断分布格局的准确性、有效性和可行性方面优于空间点格局分析[26]。
林分密度是预测林分动态的重要空间信息之一[27]。以往研究表明,种群分布格局对密度具有一定的依赖性,密度越大,聚集强度越高[28]。在本研究中,闭花木的密度最大,聚集程度也是最高的。尽管苹婆与鱼骨木在数量上存在显著差异,但它们在聚集体中所占的比例却几乎相同。这可能是在北热带喀斯特季节性雨林中岩石的裸露率较高,在岩石裸露度高的区域,植物聚集度就会偏低,反之亦然。因此,种群的分布格局与密度无明显的线性关系。
3.2 生活型对随机体的影响
植物生活型的形成是植物在特定环境中趋同适应的结果。灌木主要处于上层乔木树种的林冠下或林隙中,在数量特征和空间分布格局上会受后者的制约与影响[29]。本研究中,乔木分布比灌木多,优势度位于前10的皆为乔木树种,生活型为灌木的树种仅占25.8%。灌木树种相对于乔木树种其聚集体比例更高,分布格局为聚集分布,而乔木树种为随机分布。研究表明,不同生活型的种群聚集度差异很大,从乔木层到灌木层,聚集度逐渐增加,在林下灌丛层中,聚集度最高[25],与本文结果一致。因此随机体分布与树种自身的生物学特性有一定关系。
3.3 环境因子对随机体的影响
随机体受诸多因素的影响,不同因素的影响程度不同。本研究中,植株个体数、平均胸径、平均海拔、地形湿润指数和坡度对随机体的影响较大,而最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数对随机体的影响较小。从相关系数可以看出,植株个体数与随机体呈显著正相关,植株的个体数越多,随机体所占比例越高,群落越稳定。
林木间的竞争是最直观的相互影响关系。在林区,因为森林资源有限,为了生存,林木之间必然要进行争夺[30-32]。林木自身遗传物质的差异,导致了林木自身的竞争能力也不一样。竞争力可以从林木的胸径等属性中反映出来[33]。不同树种之间的竞争强度会影响其生长状况,从而影响其在森林中的分布。此外,强烈的竞争还会造成一些树木的死亡,从而使森林随机体的分布发生改变。本研究中优势种随机体的分布情况不一,既存在随机分布,也存在聚集分布。因此,竞争是影响随机体分布的重要原因之一。
海拔对植物的分布起重要作用,一方面是直接作用,另一方面是对其他非生物资源(如光、温、水、土等)的分配产生影响。然而,在本研究中,地形因子对于随机体分布的解释力度相对较低。尽管如此,在这些地形因子之中,海拔仍然是解释随机体分布最具影响力的因素。在北热带喀斯特季节性雨林中,物种丰富度随着海拔梯度的变化而呈现显著差异[34],物种之间的竞争强度和自疏程度都有所提高,而同种植物之间的聚集程度则有所下降[35]。同时,由于海拔的不同,水热条件也会发生较大的差异,从而使植物在不同海拔有明显的分布差异。在海拔较高的地区,如山顶,其光照强度高、水分缺乏、昼夜温差大、岩石裸露度高、土层薄。而在洼地边缘通常伴有季节性水淹,光照强度较弱[36]。因此,海拔直接或间接影响林木分布[37],造成随机体分布的差异。
4. 结论
本研究分析了北热带喀斯特季节性雨林森林动态监测样地中的随机体分布特征及其影响因素,发现该区林分整体稳定性较高,多数林木处于随机分布状态。其中,闭花木、苹婆和广西牡荆树种占主要优势,且其角尺度对林分整体分布起关键作用。不同生活型植物的随机体分布不同,树种本身生物学特性是影响不同生活型树种随机体分布的原因。不同环境因子对随机体的影响程度不同,原有植株个体数、平均胸径、平均海拔、地形湿润度指数和坡度是随机体的主要驱动因子。
-
![]()
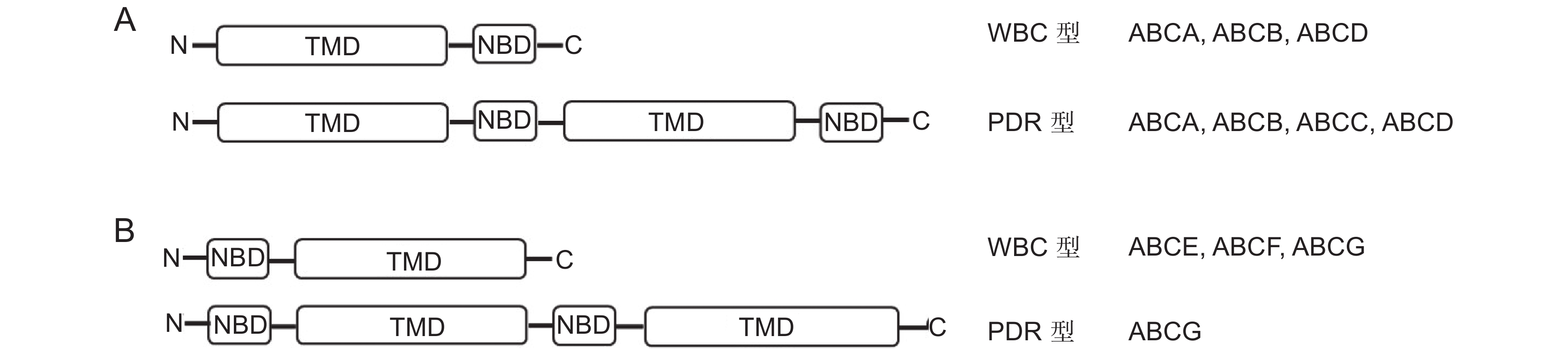
图 1 ABC转运蛋白结构
A:前向转运体结构,B:反向转运体结构。
Figure 1. Structure of ABC transporter protein
A: Forward transporter structure, B: Reverse transporter structure.
![]()
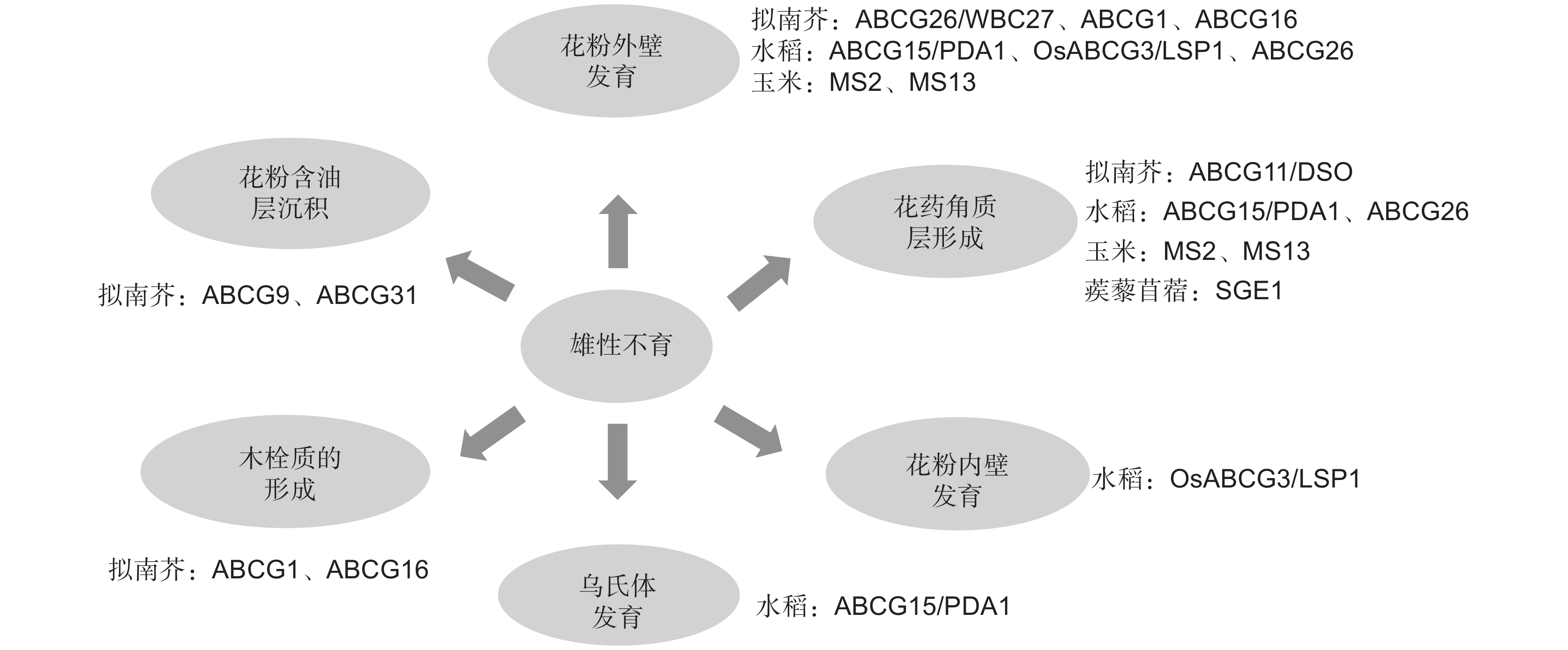
图 2 ABCG转运蛋白参与植物雄性育性调控的具体功能
Figure 2. Specific functions of ABCG transporters involved in male fertility regulation in plants
表 1 参与植物雄性育性调控的ABCG转运蛋白
Table 1 ABCG transporters involved in regulation of plant male fertility
物种
Species基因名称
Gene name转运蛋白类型
Transport protein type具体功能
Functions文献
References拟南芥
Arabidopsis thaliana (L.) Heynh.ABCG26/
WBC27半分子 参与花粉外壁形成过程中多聚酮化合物的运输,该基因突变后,花粉外壁发育异常,植株育性显著下降 [25-28] 水稻Oryza sativa L. ABCG15/
PDA1半分子 参与花药角质层、乌氏体及花粉外壁发育 [29, 30] 玉米Zea mays L. MS2 半分子 参与花粉外壁发育,GC-MS分析结果显示ms2突变体花药角质和蜡质单体显著下降 [31] 拟南芥 ABCG1 半分子 参与木栓质和花粉壁细胞外屏障形成,还可影响拟南芥生殖器官中生长素的信号传导 [33, 34, 37] 拟南芥 ABCG16 半分子 参与木栓质和花粉壁细胞外屏障形成,还可影响拟南芥生殖器官中生长素的信号传导 [33, 34, 37] 水稻 OsABCG3/
LSP1半分子 参与转运绒毡层和花粉壁合成的前体物质用于植物生殖发育,该基因功能缺失后会引起花药绒毡层异常降解、花粉外壁内层及花粉内壁层缺失 [35, 36] 拟南芥 ABCG9 半分子 与ABCG31协同将特定甾醇糖苷转运到花粉表面用于花粉含油层的沉积 [38] 拟南芥 ABCG31 全分子 与ABCG9协同将特定甾醇糖苷转运到花粉表面用于花粉含油层的沉积 [38] 拟南芥 ABCG12/
CER5半分子 参与从表皮细胞内转运蜡质前体至细胞表面 [40] 拟南芥 ABCG11/
DSO半分子 转运长链脂肪酸和超长链脂肪酸 [41, 42] 水稻 ABCG26 半分子 主要负责将脂质物质从绒毡层转运到花药壁表面供角质层形成。该基因突变后,突变体花粉外壁及花药角质层发育存在缺陷,还可以影响花粉管伸长 [23, 43] 蒺藜苜蓿
Medicago truncatula Gaertn.SGE1 半分子 影响蒺藜苜蓿花器官中蜡质和角质等长链脂肪酸的运输 [44] 玉米 ZmMS13 半分子 影响胼胝质降解、绒毡层程序性死亡和花粉外壁以及花药角质层的形成 [45] 拟南芥 ABCG28 半分子 将多胺和活性氧定位于正在生长的花粉管尖端 [46]  下载: 导出CSV
下载: 导出CSV
-
[1] Chang ZY,Chen ZF,Wang N,Xie G,Lu JW,et al. Construction of a male sterility system for hybrid rice breeding and seed production using a nuclear male sterility gene[J]. Proc Natl Acad Sci USA,2016,113(49):14145−14150.
[2] Chen XY,Zhang H,Sun HY,Luo HB,Zhao L,et al. IRREGULAR POLLEN EXINE1 is a novel factor in anther cuticle and pollen exine formation[J]. Plant Physiol,2017,173(1):307−325.
[3] Ni F,Qi J,Hao QQ,Lyu B,Luo MC,et al. Wheat Ms2 encodes for an orphan protein that confers male sterility in grass species[J]. Nat Commun,2017,8:15121.
[4] Peng XQ,Wang ML,Li YQ,Yan W,Chang ZY,et al. Lectin receptor kinase OsLecRK-S.7 is required for pollen development and male fertility[J]. J Integr Plant Biol,2020,62(8):1227−1245.
[5] Zhang DB,Luo X,Zhu L. Cytological analysis and genetic control of rice anther development[J]. J Genet Genomics,2011,38(9):379−390.
[6] Kunst L,Samuels AL. Biosynthesis and secretion of plant cuticular wax[J]. Prog Lipid Res,2003,42(1):51−80.
[7] Samuels L,Kunst L,Jetter R. Sealing plant surfaces:cuticular wax formation by epidermal cells[J]. Annu Rev Plant Biol,2008,59:683−707.
[8] Ariizumi T,Toriyama K. Genetic regulation of sporopollenin synthesis and pollen exine development[J]. Annu Rev Plant Biol,2011,62(1):437−460.
[9] Bubert H,Lambert J,Steuernagel S,Ahlers F,Wiermann R. Continuous decomposition of sporopollenin from pollen of Typha angustifolia L. by acidic methanolysis[J]. Z Naturforsch C,2002,57(11-12):1035−1041.
[10] Shi JX,Cui MH,Yang L,Kim YJ,Zhang DX. Genetic and biochemical mechanisms of pollen wall development[J]. Trends Plant Sci,2015,20(11):741−753.
[11] Jiang J,Zhang Z,Cao J. Pollen wall development:the associated enzymes and metabolic pathways[J]. Plant Biol,2013,15(2):249−263.
[12] Dhara A,Raichaudhuri A. ABCG transporter proteins with beneficial activity on plants[J]. Phytochemistry,2021,184:112663.
[13] Higgins CF. ABC Transporters:from microorganisms to man[J]. Annu Rev Cell Biol,1992,8:67−113.
[14] Theodoulou FL. Plant ABC transporters[J]. Biochim Biophys Acta Biomembr,2000,1465(1-2):79−103.
[15] Tarr PT,Tarling EJ,Bojanic DD,Edwards PA,Baldán Á. Emerging new paradigms for ABCG transporters[J]. Biochim Biophys Acta Mol Cell Biol Lipids,2009,1791(7):584−593.
[16] McFarlane HE,Shin JJH,Bird DA,Samuels AL. Arabidopsis ABCG transporters,which are required for export of diverse cuticular lipids,dimerize in different combinations[J]. Plant Cell,2010,22(9):3066−3075.
[17] Verrier PJ,Bird D,Burla B,Dassa E,Forestier C,et al. Plant ABC proteins-a unified nomenclature and updated inventory[J]. Trends Plant Sci,2008,13(4):151−159.
[18] Rea PA. Plant ATP-binding cassette transporters[J]. Annu Rev Plant Biol,2007,58:347−375.
[19] Kretzschmar T,Burla B,Lee Y,Martinoia E,Nagy R. Functions of ABC transporters in plants[J]. Essays Biochem,2011,50(1):145−160.
[20] Wu SW,Fang CW,Li ZW,Wang YB,Pan SS,et al. ATP-binding cassette G transporters and their multiple roles especially for male fertility in Arabidopsis,rice and maize[J]. Int J Mol Sci,2022,23(16):9304.
[21] Kang J,Park J,Choi H,Burla B,Kretzschmar T,et al. Plant ABC transporters[J]. Arabidopsis Book,2011,9:e0153.
[22] Elejalde-Palmett C,Segundo IMS,Garroum I,Charrier L,de Bellis D, et al. ABCG transporters export cutin precursors for the formation of the plant cuticle[J]. Curr Biol,2021,31(10):2111−2123.
[23] Zhao GC,Shi JX,Liang WQ,Xue FY,Luo Q,et al. Two ATP Binding Cassette G transporters,rice ATP binding cassette G26 and ATP binding cassette G15,collaboratively regulate rice male reproduction[J]. Plant Physiol,2015,169(3):2064−2079.
[24] Zhao GC,Shi JX,Liang WQ,Zhang DB. ATP binding cassette G transporters and plant male reproduction[J]. Plant Signal Behav,2016,11(3):e1136764.
[25] Quilichini TD,Friedmann MC,Samuels AL,Douglas CJ. ATP-binding cassette transporter G26 is required for male fertility and pollen exine formation in Arabidopsis[J]. Plant Physiol,2010,154(2):678−690.
[26] Dou XY,Yang KZ,Zhang Y,Wang W,Liu XL,et al. WBC27,an adenosine tri-phosphate-binding cassette protein,controls pollen wall formation and patterning in Arabidopsis[J]. J Integr Plant Biol,2011,53(1):74−88.
[27] Choi H,Jin JY,Choi S,Hwang JU,Kim YY,et al. An ABCG/WBC-type ABC transporter is essential for transport of sporopollenin precursors for exine formation in developing pollen[J]. Plant J,2011,65(2):181−193.
[28] Quilichini TD,Samuels AL,Douglas CJ. ABCG26-mediated polyketide trafficking and hydroxycinnamoyl spermidines contribute to pollen wall exine formation in Arabidopsis[J]. Plant Cell,2014,26(11):4483−4498.
[29] Qin P,Tu B,Wang YP,Deng LC,Quilichini TD, et al. ABCG15 encodes an ABC transporter protein,and is essential for post-meiotic anther and pollen exine development in rice[J]. Plant Cell Physiol,2013,54(1):138−154.
[30] Zhu L,Shi JX,Zhao GC,Zhang DB,Liang WQ. Post- meiotic deficient anther1 (PDA1) encodes an ABC transporter required for the development of anther cuticle and pollen exine in rice[J]. J Plant Biol,2013,56(1):59−68.
[31] Xu QL,Yang L,Kang D,Ren ZJ,Liu YJ. Maize MS2 encodes an ATP-binding cassette transporter that is essential for anther development[J]. Crop J,2021,9(6):1301−1308.
[32] Xu J,Yang CY,Yuan Z,Zhang DS,Gondwe MY,et al. The ABORTED MICROSPORES regulatory network is required for postmeiotic male reproductive development in Arabidopsis thaliana[J]. Plant Cell,2010,22(1):91−107.
[33] Yadav V,Molina I,Ranathunge K,Castillo IQ,Rothstein SJ,Reed JW. ABCG transporters are required for suberin and pollen wall extracellular barriers in Arabidopsis[J]. Plant Cell,2014,26(9):3569−3588.
[34] Yim S,Khare D,Kang J,Hwang JU,Liang WQ,et al. Postmeiotic development of pollen surface layers requires two Arabidopsis ABCG-type transporters[J]. Plant Cell Rep,2016,35(9):1863−1873.
[35] Chang ZY,Jin MN,Yan W,Chen H,Qiu SJ,Fu S,et al. The ATP-binding cassette (ABC) transporter OsABCG3 is essential for pollen development in rice[J]. Rice,2018,11(1):58.
[36] Luo T,Zou T,Yuan GQ,He ZY,Li WJ, et al. Less and shrunken pollen 1(LSP1) encodes a member of the ABC transporter family required for pollen wall development in rice(Oryza sativa L. ) [J]. Crop J,2019,8(3):492−504.
[37] Liu LP,Zhao LH,Chen PJ,Cai HY,Hou ZM,et al. ATP binding cassette transporters ABCG1 and ABCG16 affect reproductive development via auxin signalling in Arabidopsis[J]. Plant J,2020,102(6):1172−1186.
[38] Choi H,Ohyama K,Kim YY,Jin JY,Lee SB,et al. The role of Arabidopsis ABCG9 and ABCG31 ATP binding cassette transporters in pollen fitness and the deposition of steryl glycosides on the pollen coat[J]. Plant Cell,2014,26(1):310−324.
[39] Yang XJ,Wu D,Shi JX,He Y,Pinot F,et al. Rice CYP703A3,a cytochrome P450 hydroxylase,is essential for development of anther cuticle and pollen exine[J]. J Integr Plant Biol,2014,56(10):979−994.
[40] Pighin JA,Zheng HQ,Balakshin LJ,Goodman IP,Western TL,et al. Plant cuticular lipid export requires an ABC transporter[J]. Science,2004,306(5696):702−704.
[41] Bird D,Beisson F,Brigham A,Shin J,Greer S,et al. Characterization of Arabidopsis ABCG11/WBC11,an ATP binding cassette (ABC) transporter that is required for cuticular lipid secretion[J]. Plant J,2007,52(3):485−498.
[42] Panikashvili D,Shi JX,Bocobza S,Franke RB,Schreiber L,et al. The Arabidopsis DSO/ABCG11 transporter affects cutin metabolism in reproductive organs and suberin in roots[J]. Mol Plant,2010,3(3):563−575.
[43] Chang ZY,Chen ZF,Yan W,Xie G,Lu JW,et al. An ABC transporter,OsABCG26,is required for anther cuticle and pollen exine formation and pollen-pistil interactions in rice[J]. Plant Sci,2016,253:21−30.
[44] Zhu BT,Li H,Xia XZ,Meng YY,Wang N,et al. ATP-binding cassette G transporters SGE1 and MtABCG13 control stigma exsertion[J]. Plant Physiol,2020,184(1):223−235.
[45] Fang CW,Wu SW,Niu CF,Hou QC,An XL,et al. Triphasic regulation of ZmMs13 encoding an ABCG transporter is sequentially required for callose dissolution,pollen exine and anther cuticle formation in maize[J]. J Adv Res,2022,49:15−30.
[46] Do THT,Choi H,Palmgren M,Martinoia E,Hwang JU,Lee Y. Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes[J]. Proc Natl Acad Sci USA,2019,116(25):12540−12549.
-
期刊类型引用(11)
1. 王世彤,宋帅帅,李杰华,杨腾,何艺琴,魏新增,江明喜. 极小种群野生植物黄梅秤锤树的光合生理特性. 生态学杂志. 2024(03): 701-708 .  百度学术
百度学术
2. 魏新增,蒲云海,史红文,肖之强,江明喜. 湖北省国家重点保护野生植物分布与研究进展. 广西植物. 2024(11): 2000-2009 . 百度学术
3. 常二梅,刘建锋,黄跃宁,李红丽,单冰燕,江泽平,赵秀莲. 岷江柏野生居群和迁地保护居群的遗传多样性比较. 植物研究. 2022(05): 772-779 . 百度学术
4. 许玥,臧润国. 中国极小种群野生植物保护理论与实践研究进展. 生物多样性. 2022(10): 84-105 . 百度学术
5. 江全,郑旭,张康,展阳,唐罗忠. 秤锤树的种实基本性状与种子休眠机理研究. 西南林业大学学报(自然科学). 2021(02): 145-150 . 百度学术
6. 谢春平,刘大伟,吴显坤,薛晓明,南程慧. 基于灰色关联度分析的浙江楠在江苏的适宜引种地评估. 云南农业大学学报(自然科学). 2021(02): 330-337 . 百度学术
7. 黄成名,李方俊,苏彩晴,张海玲,李林. 湖北省秤锤树保护性引种试验. 甘肃林业科技. 2020(02): 5-6+27 . 百度学术
8. 杨腾,王世彤,魏新增,江明喜. 中国特有属秤锤树属植物的潜在分布区预测. 植物科学学报. 2020(05): 627-635 . 本站查看
9. 商侃侃,张希金,宋坤. 上海辰山植物园不同生活型木本植物枝叶大小关系的比较. 植物研究. 2020(05): 641-647 . 百度学术
10. 张丽,杨小波,农寿千,李东海,李苑菱,宋佳昱. 两种不同保护模式下坡垒种群发育特征. 生态学报. 2019(10): 3740-3748 . 百度学术
11. 索南邓登,陈卫东,林鹏程. 青藏高原野生濒危药用植物掌裂兰的生境及濒危因素分析. 广西植物. 2019(09): 1166-1179 . 百度学术
其他类型引用(3)












计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
- 被引次数: 14
