
Effect of sex and leaf shape on gas exchange parameters and chlorophyll fluorescence characteristics in Sabina chinensis (L.) Ant
-
摘要:
为分析性别及叶形对光合能力、能量分配与耗散的影响,本研究以圆柏(Sabina chinensis (L.) Ant.)雌株、雄株和雌雄同株3种个体的鳞叶和刺叶为材料,测定了气体交换参数、叶绿素荧光特性参数和非结构性碳水化合物(NSC)的含量。结果显示:雌株和雄株刺叶的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、水分利用率(WUE)和NSC显著高于鳞叶;雄株、雌株刺叶的非光化光淬灭系数(NPQ)和光能捕获能力(1/Fo−1/Fm)显著高于鳞叶,最大光化学量子效率(Fv/Fm)、实际电子传递速率(ETR)、实际光化学量子效率(ФPSⅡ)、光化光淬灭系数(qP)、潜在最大电子传递速率(rETRmax)和半饱和光强(Ik)显著低于鳞叶。雌株的Pn、NSC、NPQ和1/Fo−1/Fm显著高于雄株, Fv/Fm、ETR、qP、rETRmax和Ik显著低于雄株。相关性分析表明,NSC与Pn和1/Fo−1/Fm呈极显著正相关,与ETR呈显著负相关。研究结果表明,圆柏刺叶通过提高光能捕获能力及热散耗能力适应强光照环境,而鳞叶通过提高光能利用效率和光保护机制,能够适应弱光照环境。雌株可通过较强的光合速率获取较多的NSC,从而满足其生长繁殖的需求。
Abstract:This study investigated the effects of sex and leaf morphology on photosynthetic capacity, energy allocation, and dissipation in Sabina chinensis (L.) Ant. The experimental materials included prickly and scaly leaves from female, male, and monoecious plants. Gas exchange parameters, chlorophyll fluorescence characteristics, and non-structural carbohydrate (NSC) content were measured. Results showed that prickly leaves from both female and male plants exhibited significantly higher photosynthetic rate (Pn), the transpiration rate (Tr), the stomatal conductance (Gs), water use efficiency (WUE), and NSC compared to scaly leaves. Similarly, the non-photochemical quench (NPQ) and the ability of PSⅡ reaction center to trap energy from antenna pigment (1/Fo−1/Fm) were significantly higher in male and female prickly leaves, while maximum photochemical efficiency (Fv/Fm), the electron transfer rate (ETR), actual photochemical quantum efficiency (ФPSⅡ), photochemical quenching (qP), the highest maximum relative electron transfer rate (rETRmax), and half-saturated light intensity (Ik) were significantly lower in male and female prickly leaves compared to scaly leaves. Among sexes, female plants demonstrated significantly higher Pn, NSC, NPQ, and 1/Fo−1/Fm than male plants, while Fv/Fm, ETR, qP, rETRmax, and Ik were significantly lower in females. Correlation analysis showed a strong positive correlation between NSC and both Pn and 1/Fo−1/Fm, but a strong negative correlation between NSC and ETR. In conclusion, prickly leaves adapt to high-light environments by improving light energy capture ability and heat dissipation ability, whereas scaly leaves adapt to low-light environments by improving light energy utilization efficiency and light protection mechanisms. The coexistence of prickly and scaly leaves allows S. chinensis to meet the demands for photosynthate production and adapt to varying light conditions during growth and development. Female plants, through higher Pn, achieve greater NSC content, supporting the requirements for reproduction and growth.
-
森林空间结构是根据林木的空间位置和属性,来描述林木间相互关系的空间排列方式。而林木个体的空间分布状态,即分布格局,是研究林木在水平空间相互关系的重要指标,同时也是空间结构的基本特征之一[1]。分布格局的研究方法主要有样方法和距离法,但两种方法均存在局限性。
1999年,有学者提出了由参照树和相邻木组成的描述林分空间结构单元的方法[2]。该方法把林区内任意一棵树和距其最近的4棵相邻树组成的结构小组称为林分空间结构单元。角尺度(Wi)就是基于林分空间结构单元所构建的一个参数,用于描述林木的分布格局[3]。角尺度法主要是通过判断由参照树和相邻木构成的夹角大小,来判断相邻木围绕参照树的均匀性。该方法既可以通过图形来判断分布格局,也可以通过数值来判断,其操作方便,计算简单[4]。林木分布格局在林木和森林的生长演替过程中均发挥直接作用。因此,通过分析森林林分整体的角尺度分布特征,能有效地确定林分演替阶段,了解林分的稳定性。同时,对优势种分布格局进行分析,还能了解优势种与其他物种的关系,如竞争、共生、互助等。
随机木是指Wi=0.50的林木,随机木对应的结构单元称为随机体。该概念是Zhang和Hui[5]在2021年基于角尺度参数提出的。随机木在反映林分整体树种组成和竞争情况等方面具有重要作用。研究发现,随机体比例在天然林中不受地域分布、树种组成、林分结构、格局类型和树木竞争的影响,并由此提出了“随机体-稳定性”假说[6]。但在喀斯特区域,因其特有的“富钙、干旱、瘠薄”等异质生境,加之喀斯特复杂的地质地貌和环境因素,使其成为环境异质性与生物物种多样性的独特响应区。因此,生境异质性强的喀斯特地区天然林中,“随机体-稳定性”假说是否仍然成立,还有待检验。Li等[7]分析了喀斯特地区土壤和岩石两种立地上,不同径阶树木群体以及每种立地上两种生活型树木的随机体分布特征,结果发现随机体的比例和分布受生活型和生境异质性的影响,而与树的大小无关。然而,先前对于随机体在喀斯特地区影响因素的分析,均是以尺度较小的样地作为研究对象,在更大的森林样地尺度中,生活型是否对随机体有影响还不清楚。生境异质性的产生主要有两个原因,一是组成生境的能量和物质在时间或空间上的差异,如生物因素、水分状况等[8],二是区域生态系统运动发展的不平衡性[9]。而不同的环境因子在喀斯特地区中对随机体的影响还未知。此外,立地条件对于林木生长也是一个重要因素[10]。因此,研究喀斯特生境中不同因子对随机体的影响差异具有重要意义。
我国桂西南地区是北热带喀斯特季节性雨林的重要分布区之一,不但包含了从山顶到山坡到洼地的各种地形要素,还包含了水平方向上的高异质性以及垂直方向上的多层性,形成了一系列典型的“峰丛-洼地”生境类型[11, 12]。在喀斯特岩溶山地的地形地貌中,存在着复杂多样的小生境,这些小生境受不同因素的影响而呈现出多样性。此外,在不同的地段、坡向和坡位上,小生境的资源分配和植物组成等方面均存在明显差异。本文基于弄岗喀斯特季节性雨林15-hm2样地的第2次复查数据,从林分整体和优势种角尺度分布特征,以及生活型、生物因子和地形因子对随机体的影响进行分析。研究旨在探讨:(1)基于北热带喀斯特季节性雨林相邻木林分整体和优势种的随机体分布特征;(2)不同生活型树种在空间尺度较大的喀斯特生境中对随机体是否有影响;(3)北热带喀斯特季节性雨林不同环境因子对随机体的影响差异。
1. 材料与方法
1.1 研究区概况
弄岗国家级自然保护区地处广西龙州县与宁明县的交界地带,分弄岗、隆呼、陇瑞3个区,总面积为10 077 hm2。保护区土壤类型为原始石灰土、棕色石灰土、黑色石灰土等。年降水集中在5-9月,降水量分布不均匀,干湿交替较强;年平均温度22 ℃,最低温度13 ℃,最高温度37 ℃~39 ℃。
1.2 样地设置与调查
北热带喀斯特季节性雨林地处弄岗自然保护区弄岗片的弄姆皇(22°25′N,106°57′E)。整个样地南北宽300 m,东西长500 m,海拔为180~370 m,平均海拔260 m,样地坡度范围在3.7°~78.9°,平均坡度41.7°。整个样地生境多样化,属于喀斯特“峰丛-洼地”典型的生境类型,在中国森林生物多样性监测网络中,该样地是唯一的典型热带喀斯特森林监测样地。
根据CTFS全球森林生物多样性监测标准,利用全站仪将15-hm2样地划分成1 500个10 m×10 m的样方。在此基础上,于2011年完成了首次植被调查,对样方内胸径(DBH)≥1 cm个体的名称、胸径、坐标及生长状态等进行了调查并记录。之后,每5年进行一次调查,并于2021年完成第2次复查。本文以2021年的复查数据为依据,选取DBH≥5 cm的个体作为研究对象。
1.3 树种优势度的计算
树种优势度(Dsp)指树种在林分中的数量优势度和空间上的优势度,选取优势度位于前10的物种作为优势种,Dsp的计算公式为:
Dsp=√Dg⋅(1−¯Usp) (1) 式中,Dg表示相对显著度,
ˉUsp 表示树种大小比数均值。Dsp接近1表示非常优势,接近0表示几乎没有优势[13]。树种大小比数均值表达式如下:
¯Usp=1Nsp∑Nspi=1Ui (2) 式中,Nsp表示所观察树种的参照树数量,Ui为树种的第i株树的大小比数值。
1.4 角尺度计算
角尺度(Wi)指的是参照树与4株最近邻木间小于标准角的夹角占比,用下式表示[3]:
Wi=1n∑nj=1Zij (3) 其中:Zij=
1{1(当j个角小于标准角α0)0(否则) 式中,Wi为参照树i的角尺度,n指相邻木个体数,n=4。Zij为参照树i与其相邻木j之间的第j个夹角α与标准角α0之间的大小关系,有两种可能的值,当Zij=1时,表明第j个α角小于标准角,当Zij=0时,表明第j个α角大于标准角。
角尺度均值(
¯W )能够反映一个林分整体的分布情况。其计算公式如下:¯W=1N∑Niwi (4) 优势种角尺度均值用下式表示:
¯Wsp=1Nsp∑Nspi=1 (5) 式中,N为样地林木总株数。当角尺度均值在[0.457,0.517]时为随机分布;大于0.517为聚集分布;小于0.475为均匀分布[14]。角尺度等于0.50的林木被称为随机木,相应林木的结构单元称为随机体[15]。角尺度为0.75或1的林木为聚集木,相应林木结构单元称为聚集体。角尺度为0或0.25的林木被称作均匀木,相应林木的结构单元被称作均匀体。本研究在计算林分整体和优势种角尺度时,设置了5 m缓冲区,并将缓冲区内的林木做相邻木处理,以此消除林木的边缘效应造成的影响。
1.5 Pearson 相关分析
运用Pearson相关分析法,判断随机体与环境因子及环境因子间的相关性。相关系数r计算公式如下:
r=∑(x−ˉx)−(y−ˉy)∑(x−ˉx)2∑(y−ˉy)2 (6) r取值范围为−1~1,当|r|≥0.8,表示两者高度相关;0.5≤|r|<0.8,为中度相关;0.3≤|r|<0.5,表示低相关;当|r|<0.3,表示相关性非常微弱,可视为没有相关性,P<0.05表明有统计学意义。
1.6 环境因子获取
用全站仪获取精确海拔数据,计算每个20 m×20 m样方的平均海拔、坡向、坡度和凹凸度4个地形指标[16-18]。土壤湿度状况以地形湿润指数和干旱度指数为表征,参见Punchi-Manage等[19]的研究。样地内岩石裸露情况采用踏查法进行评估。综上共有7个地形因子。此外,还考查了每个样方内最大胸径、胸高断面积之和、植株个体数和平均胸径4个生物因子,共11个环境因子。
1.7 冗余分析
冗余分析的响应变量由每个样方的随机体个数组成。为了消除林木边缘效应的影响,设置2 m缓冲区,缓冲区内的林木做相邻木处理。解释变量数据包括11个环境因子。冗余分析前,对数据进行标准化处理,并比较自变量的相关系数是否大于7,以此衡量自变量间的共线性。再计算每个变量的方差膨胀因子,当方差膨胀因子<10时,表示无明显的共线性问题[20],可进行冗余分析。采用层次分割方法对冗余分析结果进行变差分解,以获取每个解释变量的校正解释率。
1.8 数据处理
数据分析计算均利用R 4.3.1软件完成,冗余分析在vegan程序包中进行,层次分割采用rdacca.hp程序包[21],利用ForestSAS程序包计算角尺度和大小比数均值[22],Pearson相关系数图在corrplot包中绘制,角尺度分布特征图在ggplot2包中完成,单独效应比例图在Origin 2021软件中绘制。
2. 结果与分析
2.1 林分整体和优势种随机体的比例和分布特征
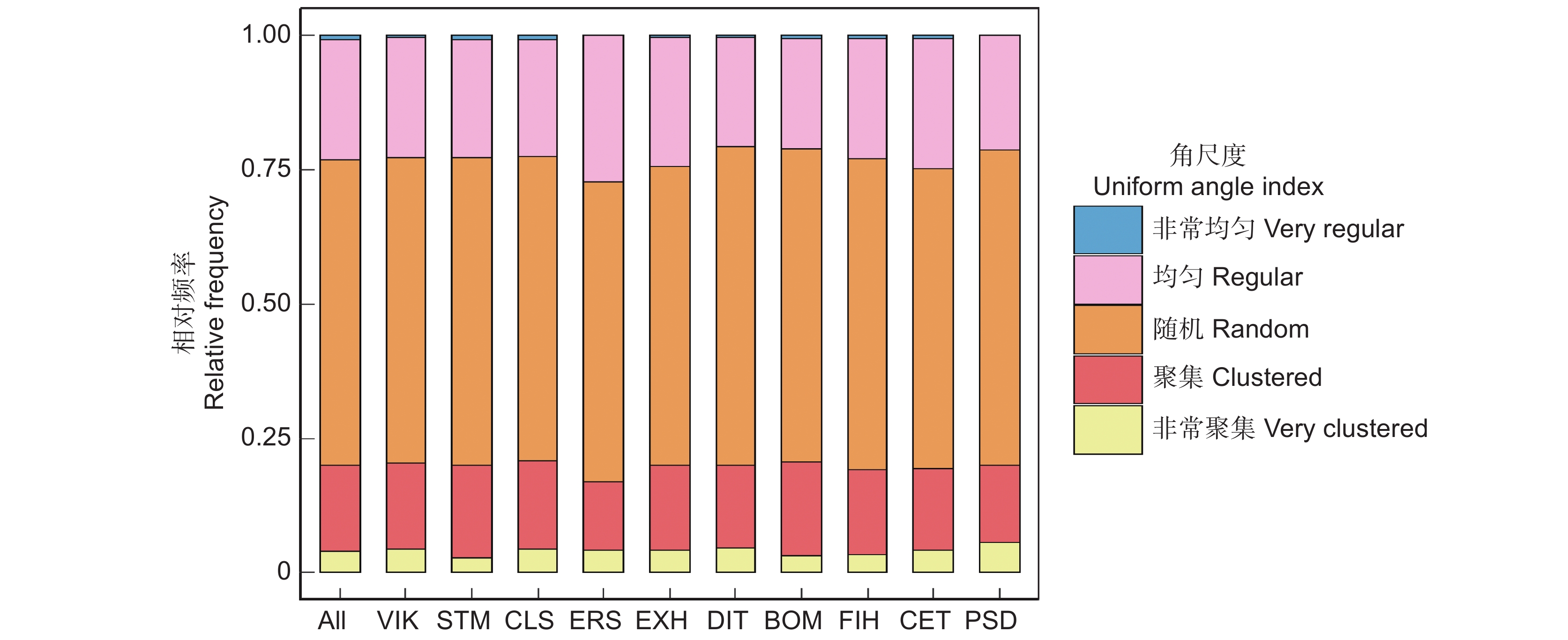
树种优势度位于前10的物种在森林中起关键作用,尤其是闭花木(Cleistanthus sumatranus (Miq.) Muell. Arg.)、苹婆(Sterculia monosperma Ventenat)和广西牡荆(Vitex kwangsiensis P'ei),不仅在数量上占林分整体的绝大多数,其优势度也是最高的,分别为0.262、0.260、0.245,其他树种的优势度均小于0.200。角尺度计算结果表明,林分整体和优势种角尺度均值都在随机分布范围内(表1)。但林分整体和优势种随机木的分布频率均超过0.5,而均匀木和聚集木分布频率均比随机木低,说明样地中大多数林木的最近相邻木处于随机分布,各个优势种的最近相邻木处于随机分布的也占大多数,即随机体占绝对分布(图1)。
表 1 北热带喀斯特季节性雨林树种优势度排名前10的树种Table 1. Top 10 dominant species in northern tropical karst seasonal rainforest树种
Species科名
Family株数
Number of
plants大小比数
Dominance相对显著度
Relative
significance优势度
Advantage
degree角尺度
Uniform angle
index广西牡荆
Vitex kwangsiensis P'ei唇形科 1 645 0.325 0.122 0.286 0.504 苹婆
Sterculia monosperma Ventenat梧桐科 2 426 0.491 0.134 0.262 0.498 闭花木
Cleistanthus sumatranus (Miq.) Muell. Arg.大戟科 3 054 0.530 0.139 0.256 0.504 劲直刺桐
Erythrina stricta Roxb.豆科 242 0.074 0.031 0.171 0.485 蚬木
Excentrodendron tonkinense (A. Chev.)
H. T. Chang et R. H. Miau锦葵科 483 0.367 0.039 0.156 0.498 海南椴
Diplodiscus trichosperma (Merrill) Y. Tang锦葵科 456 0.317 0.033 0.150 0.509 黄梨木
Boniodendron minus (Hemsl.) T. Chen无患子科 404 0.314 0.026 0.133 0.506 对叶榕
Ficus hispida L. f.桑科 676 0.602 0.031 0.110 0.497 假玉桂
Celtis timorensis Span大麻科 414 0.509 0.020 0.099 0.495 鱼骨木
Psydrax dicocca Gaertn.茜草科 252 0.405 0.015 0.094 0.510 林分整体 − 10 052 0.489 − − 0.500 ![]() 图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.
图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.2.2 不同生活型树种随机体的比例和分布特征
乔木角尺度均值在随机分布范围内,为0.499,随机体占比为57.167%,均匀体占比较小,为23.189%,聚集体次之,为19.644%。灌木角尺度均值为0.519,为聚集分布,随机体占比为59.173%,均匀体占比为19.064%,聚集体为21.763%。乔木和灌木的随机体比例均超过55%,但灌木的聚集体占比高于均匀体。而在乔木中,均匀体占比高于聚集体,表明不同生活型的植物,其随机体比例和分布存在一定的差异(表2)。
表 2 北热带喀斯特季节性雨林不同生活型随机体比例和分布Table 2. Proportion and distribution of random framework among different life forms in northern tropical karst seasonal rainforest生活型
Life style角尺度 Uniform angle index / % 非常均匀
Very regular均匀
Regular随机
Random聚集
Clustered非常聚集
Very clustered角尺度均值
Uniform angle index mean乔木 0.773 22.416 57.167 15.785 3.859 0.499 灌木 0.719 18.345 59.173 16.187 5.576 0.519 2.3 Pearson相关性分析
研究结果显示,随机体与胸高断面积之和、平均海拔、坡度、岩石裸露率、凹凸度、干旱度指数、原有植株个体数之间均存在显著正相关,其中相关性最高的为原有植株个体数,相关系数达0.86;平均海拔和坡度分别与随机体存在低度相关,相关系数分别为0.36和0.31。随机体分别与最大胸径、平均胸径和地形湿润指数存在显著负相关,但与坡向无显著相关性。胸高断面积和最大胸径、坡度和地形湿润指数间也具有较高的相关性,相关系数分别为0.7和−0.72(图2)。
![]() 图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.
图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.2.4 环境因子与随机体的冗余分析及单个解释变量的相对重要性
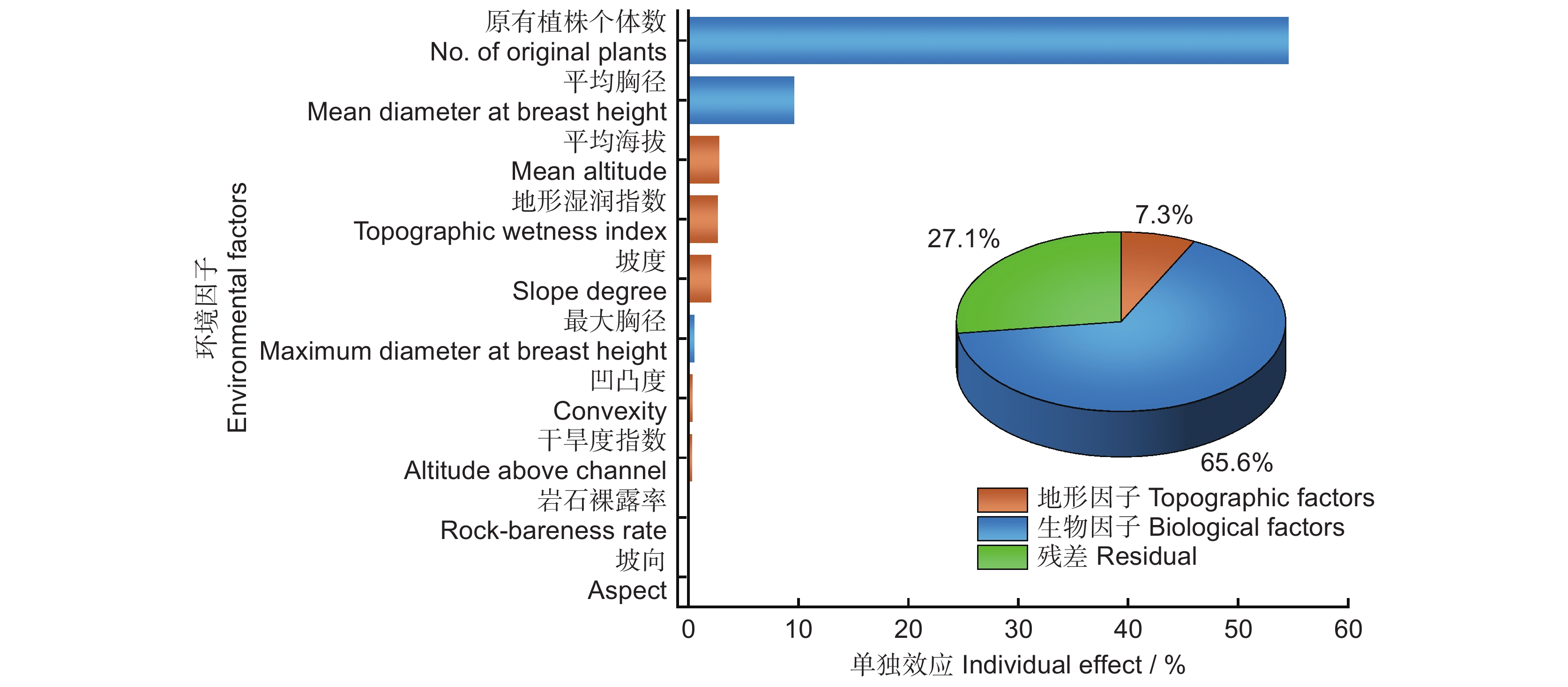
结合环境因子的相关性和方差膨胀因子分析,将胸高断面积之和剔除。对随机体与生物因子和地形因子进行冗余分析,10个环境因子对随机体的解释量R2经校正后为0.729。其中,生物因子解释了65.6%,地形因子解释了7.3%。未被解释的部分占27.1%。根据层次分割法各环境因子单独效应比例排序,随机体的主要影响因素包括原有植株个体数、平均胸径、平均海拔、地形湿润指数和坡度;贡献较小的是最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数(图3)。
![]() 图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest
图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest3. 讨论
3.1 随机体的分布特征
林木的空间分布格局主要分为随机分布、规则分布和聚集分布,最理想的是随机分布,而随机分布也是天然原始顶极群落中最为显著的结构特征[23]。本研究中,林分整体为随机分布,优势种的角尺度均值也在随机分布范围内,且随机体占比均超过50%。因此,随机体以相似的比例出现在不同树种中,且支持了“随机体-稳定性”假说,与前人研究结果一致[5]。这种结构体相对来说具有较小的生存压力,稳定性高,在竞争情况下不易出现弱势或者不健康的林木,在自然演替过程中存活几率更大[6]。而均匀体和聚集体占比较小,与袁星明等[24]的报道相似。
不同空间分布格局分析方法可能导致结果上的差异。前人研究采用完全随机零模型和泊松异质性零模型对该地区分布格局进行了分析,完全随机零模型结果表明,在小尺度上所有物种均呈聚集分布[25],与本研究结果不一致;然而,当泊松异质性零模型去除生境异质性后,大部分物种表现为随机分布,与本文结果一致。完全随机零模型是基于假设个体随机分布,不受任何因素的干扰而对分布格局进行拟合分析。但在生境异质性强的北热带喀斯特地区,物种的分布显然会受到各种因素的影响。因此,在不考虑任何因素干扰情况下大多数物种呈聚集分布,与本研究结果存在差异。泊松异质性零模型主要用于模拟生境关联,本研究基于相邻木的角尺度法分析林木分布格局,也未直接考虑生境异质性,所以二者结果一致。另外,角尺度法不存在尺度效应,而空间点格局法可以拟合分析不同尺度的林木分布格局,二者各具特点。对于这两种方法,有学者也进行了对比,发现角尺度法在判断分布格局的准确性、有效性和可行性方面优于空间点格局分析[26]。
林分密度是预测林分动态的重要空间信息之一[27]。以往研究表明,种群分布格局对密度具有一定的依赖性,密度越大,聚集强度越高[28]。在本研究中,闭花木的密度最大,聚集程度也是最高的。尽管苹婆与鱼骨木在数量上存在显著差异,但它们在聚集体中所占的比例却几乎相同。这可能是在北热带喀斯特季节性雨林中岩石的裸露率较高,在岩石裸露度高的区域,植物聚集度就会偏低,反之亦然。因此,种群的分布格局与密度无明显的线性关系。
3.2 生活型对随机体的影响
植物生活型的形成是植物在特定环境中趋同适应的结果。灌木主要处于上层乔木树种的林冠下或林隙中,在数量特征和空间分布格局上会受后者的制约与影响[29]。本研究中,乔木分布比灌木多,优势度位于前10的皆为乔木树种,生活型为灌木的树种仅占25.8%。灌木树种相对于乔木树种其聚集体比例更高,分布格局为聚集分布,而乔木树种为随机分布。研究表明,不同生活型的种群聚集度差异很大,从乔木层到灌木层,聚集度逐渐增加,在林下灌丛层中,聚集度最高[25],与本文结果一致。因此随机体分布与树种自身的生物学特性有一定关系。
3.3 环境因子对随机体的影响
随机体受诸多因素的影响,不同因素的影响程度不同。本研究中,植株个体数、平均胸径、平均海拔、地形湿润指数和坡度对随机体的影响较大,而最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数对随机体的影响较小。从相关系数可以看出,植株个体数与随机体呈显著正相关,植株的个体数越多,随机体所占比例越高,群落越稳定。
林木间的竞争是最直观的相互影响关系。在林区,因为森林资源有限,为了生存,林木之间必然要进行争夺[30-32]。林木自身遗传物质的差异,导致了林木自身的竞争能力也不一样。竞争力可以从林木的胸径等属性中反映出来[33]。不同树种之间的竞争强度会影响其生长状况,从而影响其在森林中的分布。此外,强烈的竞争还会造成一些树木的死亡,从而使森林随机体的分布发生改变。本研究中优势种随机体的分布情况不一,既存在随机分布,也存在聚集分布。因此,竞争是影响随机体分布的重要原因之一。
海拔对植物的分布起重要作用,一方面是直接作用,另一方面是对其他非生物资源(如光、温、水、土等)的分配产生影响。然而,在本研究中,地形因子对于随机体分布的解释力度相对较低。尽管如此,在这些地形因子之中,海拔仍然是解释随机体分布最具影响力的因素。在北热带喀斯特季节性雨林中,物种丰富度随着海拔梯度的变化而呈现显著差异[34],物种之间的竞争强度和自疏程度都有所提高,而同种植物之间的聚集程度则有所下降[35]。同时,由于海拔的不同,水热条件也会发生较大的差异,从而使植物在不同海拔有明显的分布差异。在海拔较高的地区,如山顶,其光照强度高、水分缺乏、昼夜温差大、岩石裸露度高、土层薄。而在洼地边缘通常伴有季节性水淹,光照强度较弱[36]。因此,海拔直接或间接影响林木分布[37],造成随机体分布的差异。
4. 结论
本研究分析了北热带喀斯特季节性雨林森林动态监测样地中的随机体分布特征及其影响因素,发现该区林分整体稳定性较高,多数林木处于随机分布状态。其中,闭花木、苹婆和广西牡荆树种占主要优势,且其角尺度对林分整体分布起关键作用。不同生活型植物的随机体分布不同,树种本身生物学特性是影响不同生活型树种随机体分布的原因。不同环境因子对随机体的影响程度不同,原有植株个体数、平均胸径、平均海拔、地形湿润度指数和坡度是随机体的主要驱动因子。
-
![]()
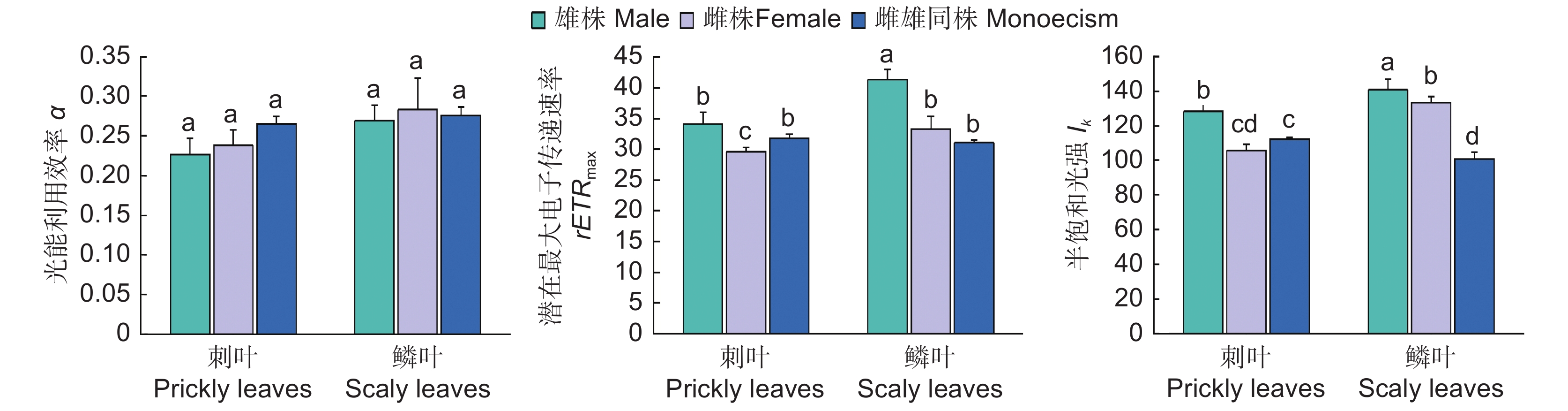
图 2 圆柏快速光响应曲线参数比较
不同小写字母表示差异显著(P<0.05)。
Figure 2. Comparison of fast light response curve parameters in Sabina chinensis
Different lowercase letters indicate significant difference (P<0.05).
表 1 4种光合参数的日变化
Table 1 Comparison of four photosynthetic parameters
参数
Parameter性别
Sex叶形
Leaf
shape时间
Time8:00 10:00 12:00 14:00 16:00 18:00 Pn / μmol·
m−2·s−1雄株 刺叶 1.2±0.10b(A) 3.5±0.10b(A) 1.7±0.10b(A) 0.4±0.02b(A) 0.9±0.20b(A) 0.1±0.10c(B) 鳞叶 0.7±0.05B(B) 2.2±0.30A(B) 1.2±0.10B(B) 0.2±0.03B(B) 0.7±0.04C(A) 0.3±0.03B(A) 雌株 刺叶 2.2±0.15a(A) 4.7±0.30a(A) 2.9±0.23a(A) 0.6±0.06a(A) 2.8±0.30a(A) 0.9±0.05a(A) 鳞叶 0.8±0.08B(B) 2.8±0.34A(B) 1.4±0.07A(B) 0.5±0.01A(B) 1.3±0.13B(B) 0.3±0.03C(B) 雌雄
同株刺叶 0.7±0.04c(B) 1.8±0.21c(B) 0.5±0.05c(B) 0.2±0.08c(B) 0.9±0.17b(B) 0.3±0.01b(B) 鳞叶 1.2±0.12A(A) 2.7±0.21A(A) 0.9±0.08C(A) 0.6±0.06A(A) 1.5±0.12A(A) 0.8±0.02A(A) Gs / mmol·
m−2·s−1雄株 刺叶 18.0±0.80a(A) 25.6±2.20a(A) 19.7±1.20b(A) 25.7±1.50a(A) 15.3±1.00b(A) 10.1±0.70a(B) 鳞叶 15.0±0.90A(B) 25.9±1.70A(A) 11.5±2.40B(B) 19.1±2.10A(B) 15.6±2.10A(A) 14.2±0.80A(A) 雌株 刺叶 19.1±1.10a(A) 22.1±1.80a(A) 24.4±1.50a(A) 17.5±1.20b(A) 29.9±1.10a(A) 10.8±0.70a(A) 鳞叶 15.3±1.00A(B) 23.1±1.40A(A) 21.1±2.10A(A) 17.7±1.20A(A) 17.8±1.50A(B) 9.8±0.99B(A) 雌雄
同株刺叶 14.0±0.90b(A) 20.1±2.10a(A) 17.0±1.60b(B) 15.0±1.30b(B) 18.0±1.80b(A) 9.9±0.56a(A) 鳞叶 14.0±0.90A(A) 23.1±3.00A(A) 23.0±1.40A(A) 21.0±1.90A(A) 19.0±1.40A(A) 11.0±0.90B(A) Tr / mmol·
m−2·s−1雄株 刺叶 0.4±0.02a(A) 1.1±0.03a(B) 1.4±0.05a(A) 1.7±0.08a(A) 0.9±0.08b(A) 0.4±0.03a(B) 鳞叶 0.3±0.05A(B) 1.2±0.06A(A) 0.8±0.04B(B) 1.0±0.03C(B) 0.7±0.04A(B) 0.7±0.09A(A) 雌株 刺叶 0.4±0.15a(A) 0.7±0.07c(A) 0.8±0.12b(B) 1.3±0.06b(B) 1.1±0.09a(A) 0.3±0.05b(B) 鳞叶 0.3±0.08A(A) 0.9±0.14B(A) 1.0±0.07A(A) 1.4±0.01A(A) 0.8±0.13A(B) 0.4±0.03B(A) 雌雄
同株刺叶 0.2±0.04b(A) 0.9±0.11b(A) 0.6±0.05c(B) 1.0±0.08c(B) 0.8±0.07b(A) 0.3±0.01b(A) 鳞叶 0.3±0.12A(A) 0.7±0.09B(A) 0.8±0.08B(A) 1.3±0.06B(A) 0.8±0.12A(A) 0.3±0.02C(A) WUE /
mmol/
mol雄株 刺叶 3.0±0.10c(A) 2.7±0.18b(A) 0.7±0.10b(B) 0.2±0.04b(A) 1.0±0.20b(A) 0.3±0.10c(B) 鳞叶 2.3±0.05C(B) 1.6±0.30C(B) 0.9±0.10A(A) 0.2±0.02B(A) 1.0±0.04C(A) 0.5±0.03B(A) 雌株 刺叶 5.5±0.15a(A) 5.3±0.30a(A) 3.5±0.23a(A) 0.5±0.06a(A) 2.5±0.30a(A) 2.7±0.05a(A) 鳞叶 2.7±0.18B(B) 2.6±0.34B(B) 1.4±0.27A(B) 0.4±0.11A(B) 1.8±0.13B(B) 0.7±0.17B(B) 雌雄
同株刺叶l 3.3±0.04b(B) 2.0±0.21c(B) 0.8±0.05b(B) 0.2±0.08b(B) 1.2±0.17b(B) 1.1±0.01b(B) 鳞叶 3.9±0.12A(A) 3.7±0.24A(A) 1.1±0.18A(A) 0.4±0.12A(A) 2.0±0.12A(A) 2.5±0.13A(A) 注:不同小写字母表示同一参数下刺叶3种性别间差异显著,不同大写字母表示同一参数下鳞叶3种性别间差异显著,括号内不同大写字母表示同一参数下各性别鳞、刺叶间差异显著(P<0.05)。 Notes: Different lowercase letters indicate significant differences between sexes of prickly leaves under the same parameter, different uppercase letters indicate significant differences between sexes of scaly leaves under the same parameter, and different uppercase letters in parentheses indicate significant differences between sexes of scaly and prickly leaves under the same parameter (P<0.05).  下载: 导出CSV
下载: 导出CSV
表 2 不同性别圆柏异形叶叶绿素荧光参数
Table 2 Chlorophyll fluorescence parameters of heterophyllous leaves of different sexes in Sabina chinensis
叶形
Leaf shape性别
Sex最大光化光效率
Fv/Fm实际电子传递速率
ETR实际光化光效率
ФPSⅡ光化学淬灭系数
qP非光化学淬灭系数
NPQ光能捕获能力
1/Fo−1/Fm刺叶 雄株 0.78±0.014b 23.60±1.248c 0.22±0.011b 0.41±0.014b 1.96±0.165a 1.71±0.145c 雌株 0.78±0.013b 21.36±1.700d 0.20±0.016b 0.37±0.015c 2.12±0.126a 1.97±0.169b 雌雄同株 0.78±0.009b 24.65±0.150c 0.23±0.001b 0.41±0.003b 1.82±0.180ab 2.35±0.198a 鳞叶 雄株 0.81±0.010a 30.44±3.440a 0.28±0.046a 0.47±0.021a 1.63±0.129b 1.58±0.164d 雌株 0.78±0.011b 27.56±1.190b 0.26±0.030ab 0.44±0.020ab 2.01±0.141a 1.63±0.092cd 雌雄同株 0.80±0.002a 24.25±0.530c 0.22±0.039b 0.42±0.036b 2.07±0.113a 2.21±0.204a 注:同列不同小写字母表示差异显著(P<0.05)。 Note: Different lowercase letters in the same column indicate significant difference (P<0.05).
下载: 导出CSV
表 3 不同叶片非结构性碳水化合物含量
Table 3 Non-structural carbohydrate content in different leaves
性别
Sex叶形
Leaf shape可溶性糖
SS / mg/g淀粉
ST / mg/g非结构性碳水化合物
NSC / mg/g雄株 刺叶 66.33±3.17aB 23.76±1.93aA 90.09±5.10aB 鳞叶 69.20±3.37aA 14.67±2.53bB 83.87±5.90bB 雌株 刺叶 74.58±2.65aA 24.92±2.06aA 99.50±4.71aA 鳞叶 65.70±3.62bAB 21.65±2.11bA 87.34±5.73bA 雌雄同株 刺叶 55.37±4.38aC 22.03±1.80aA 77.40±6.19aC 鳞叶 61.50±3.65bB 20.39±1.28aA 81.89±4.93bB 注:不同小写字母表示相同性别下不同叶形间存在显著差异;不同大写字母表示相同叶形下不同性别间存在显著差异, P<0.05。 Notes: Different lowercase letters indicate significant differences between different leaf shapes in the same sex; different capital letters indicate significant differences between different sexes with the same leaf shape, P<0.05.
下载: 导出CSV
-
[1] 李理渊,李俊,同小娟,孟平,张劲松,张静茹. 黄河小浪底栓皮栎、刺槐叶片电子传递速率-光响应的模拟[J]. 植物生态学报,2018,42(10):1009−1021. doi: 10.17521/cjpe.2018.0063 Li LY,Li J,Tong XJ,Meng P,Zhang JS,Zhang JR. Simulation on the light-response curves of electron transport rate of Quercus variabilis and Robinia pseudoacacia leaves in the Xiaolangdi area,China[J]. Chinese Journal of Plant Ecology,2018,42(10):1009−1021. doi: 10.17521/cjpe.2018.0063
[2] 王海珍,徐雅丽,代雅琦,韩路. 胡杨异形叶光合生理特性及其与叶形态的关系[J]. 塔里木大学学报,2020,32(3):35−43. doi: 10.3969/j.issn.1009-0568.2020.03.005 Wang HZ,Xu YL,Dai YQ,Han L. The photosynthetic physiology characteristics of heteromorphic leaves and their relationship with leaf shape in Populus euphratica[J]. Journal of Tarim University,2020,32(3):35−43. doi: 10.3969/j.issn.1009-0568.2020.03.005
[3] 张金玲. 臭柏对光胁迫的生理生态响应研究[D]. 保定:河北农业大学,2020:1−122. [4] Hao JQ,Yue N,Zheng CX. Analysis of changes in anatomical characteristics and physiologic features of heteromorphic leaves in a desert tree,Populus euphratica[J]. Acta Physiol Plant,2017,39(8):160. doi: 10.1007/s11738-017-2467-9
[5] 刘建丽. 常绿乔木圆柏的繁殖技术[J]. 新疆林业,2021(4):20−21. doi: 10.3969/j.issn.1005-3522.2021.04.008 [6] Tanaka-Oda A,Kenzo T,Kashimura S,Ninomiya I,Wang LH,et al. Physiological and morphological differences in the heterophylly of Sabina vulgaris Ant. in the semi-arid environment of Mu Us Desert,Inner Mongolia,China[J]. J Arid Environ,2010,74(1):43−48. doi: 10.1016/j.jaridenv.2009.07.013
[7] Zhang JL,Xu XH,Li XG,Li YL,Guy RD,Chen HP. Photoprotection in heteromorphic leaves of savin juniper (Juniperus sabina L.)[J]. Photosynthetica,2019,57(3):780−787. doi: 10.32615/ps.2019.097
[8] Zhang JL,Li XG,Xu XH,Chen HP,Li YL,Guy RD. Leaf morphology,photosynthesis and pigments change with age and light regime in savin juniper[J]. Plant Biol,2021,23(6):1097−1108. doi: 10.1111/plb.13256
[9] 王钰莹,魏梦瑶,焦梦晓,张奕蕾,邱念伟. 圆柏刺叶和鳞叶的光合功能比较[J]. 林业科技,2023,48(4):6−10,15. Wang YY,Wei MY,Jiao MX,Zhang YL,Qiu NW. Comparison of photosynthetic function between needle leaves and scale leaves of Sabina chinensis[J]. Forestry Science & Technology,2023,48(4):6−10,15.
[10] Renner SS,Ricklefs RE. Dioecy and its correlates in the flowering plants[J]. Am J Bot,1995,82(5):596−606. doi: 10.1002/j.1537-2197.1995.tb11504.x
[11] 贺俊东,胥晓,郇慧慧,杨帅,秦芳. 青杨雌雄扦插苗光合作用日变化与叶绿素荧光参数特征[J]. 植物研究,2014,34(2):219−225. He JD,Xu X,Huan HH,Yang S,Qin F. Characteristics of chlorophyll fluorescent parameters and daily dynamics of photosynthesis in female and male Populus cathayana cutting seedlings[J]. Bulletin of Botanical Research,2014,34(2):219−225.
[12] 唐学玺. 环境胁迫下雌雄异株植物的差异响应特征及研究进展[J]. 中国海洋大学学报,2020,50(7):74−81. Tang XX. Characteristics and research progress of sex-specific responses to environmental stresses of dioecious plants[J]. Periodical of Ocean University of China,2020,50(7):74−81.
[13] Zhang S,Chen LH,Duan BL,Korpelainen H,Li CY. Populus cathayana males exhibit more efficient protective mechanisms than females under drought stress[J]. For Ecol Manage,2012,275:68−78. doi: 10.1016/j.foreco.2012.03.014
[14] Xu X,Peng GQ,Wu CC,Korpelainen H,Li CY. Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana[J]. Tree Physiol,2008,28(11):1751−1759. doi: 10.1093/treephys/28.11.1751
[15] Liu M,Liu XX,Kang JY,Korpelainen H,Li CY. Are males and females of Populus cathayana differentially sensitive to Cd stress?[J]. J Hazard Mater,2020,393:122411. doi: 10.1016/j.jhazmat.2020.122411
[16] Liao J,Cai ZY,Song HF,Zhang S. Poplar males and willow females exhibit superior adaptation to nocturnal warming than the opposite sex[J]. Sci Total Environ,2020,717:137179. doi: 10.1016/j.scitotenv.2020.137179
[17] Yang F,Wang Y,Wang J,Deng WQ,Liao L,Li M. Different eco-physiological responses between male and female Populus deltoides clones to waterlogging stress[J]. For Ecol Manage,2011,262(11):1963−1971. doi: 10.1016/j.foreco.2011.08.039
[18] Li CY,Ren J,Luo JX,Lu RS. Sex-specific physiological and growth responses to water stress in Hippophae rhamnoides L. populations[J]. Acta Physiol Plant,2004,26(2):123−129. doi: 10.1007/s11738-004-0001-3
[19] 孙晓伟. 温带3种雌雄异株树种光合特征对比[D]. 哈尔滨:东北林业大学,2022:1−62. [20] Marshall JD,Dawson TE,Ehleringer JR. Gender-related differences in gas exchange are not related to host quality in the xylem-tapping mistletoe,Phoradendron juniperinum (Viscaceae)[J]. Am J Bot,1993,80(6):641−645. doi: 10.1002/j.1537-2197.1993.tb15234.x
[21] Jones MH,MacDonald SE,Henry GHR. Sex-and habitat-specific responses of a high arctic willow,Salix arctica,to experimental climate change[J]. Oikos,1999,87(1):129−138. doi: 10.2307/3547004
[22] Dawson TE,Ward JK,Ehleringer JR. Temporal scaling of physiological responses from gas exchange to tree rings:a gender-specific study of Acer negundo (Boxelder) growing under different conditions[J]. Funct Ecol,2004,18(2):212−222. doi: 10.1111/j.0269-8463.2004.00838.x
[23] Letts MG,Phelan CA,Johnson DRE,Rood SB. Seasonal photosynthetic gas exchange and leaf reflectance characteristics of male and female cottonwoods in a riparian woodland[J]. Tree Physiol,2008,28(7):1037−1048. doi: 10.1093/treephys/28.7.1037
[24] Tozawa M,Ueno N,Seiwa K. Compensatory mechanisms for reproductive costs in the dioecious tree Salix integra[J]. Botany,2009,87(3):315−323. doi: 10.1139/B08-125
[25] 马文宝,廖成云,姬慧娟,陈雪,董廷发. 濒危连香树种群性比和雌雄株功能性状的差异[J]. 生态学杂志,2019,38(8):2414−2419. Ma WB,Liao CY,Ji HJ,Chen X,Dong TF. Sex ratio and sexual difference of functional traits in the endangered plant Cercidiphyllum japonicum[J]. Chinese Journal of Ecology,2019,38(8):2414−2419.
[26] 黄云浩,辛本花,王娟. 雌雄异株植物鼠李生殖分配与生殖耗费补偿机制[J]. 北京林业大学学报,2019,41(11):31−36. Huang YH,Xin BH,Wang J. Reproductive allocation and compensation mechanism for reproductive costs of dioecious shrub Rhamnus davurica[J]. Journal of Beijing Forestry University,2019,41(11):31−36.
[27] 陈银萍. 圆柏属植物抗冷冻适应性机制研究[D]. 兰州:兰州大学,2006:1−90. [28] 杨宗娟. 两种圆柏属植物叶片抗氧化系统对低温胁迫的响应机制研究[D]. 兰州:兰州交通大学,2013:1−69. [29] 路通. 圆柏叶挥发性有机物动态变化研究[D]. 晋中:山西农业大学,2022:1−58. [30] 宋晨慧,张有福,王梦可,王露露,陈春艳. 3种圆柏属植物叶片气孔特征对自然光强的响应[J]. 广东农业科学,2022,49(4):28−34. Song CH,Zhang YF,Wang MK,Wang LL,Chen CY. Response of leaf stomatal characteristics of three species of Juniperus to natural light intensity[J]. Guangdong Agricultural Sciences,2022,49(4):28−34.
[31] 韩兴华. 4种针叶树光合蒸腾特性的研究[D]. 呼和浩特:内蒙古农业大学,2007:1−39. [32] Ralph PJ,Gademann R. Rapid light curves:a powerful tool to assess photosynthetic activity[J]. Aquat Bot,2005,82(3):222−237. doi: 10.1016/j.aquabot.2005.02.006
[33] 冀宗琪. 蒙古栎非结构性碳水化合物在干旱胁迫中的调节作用[D]. 沈阳:沈阳农业大学,2022:1−60. [34] 段娜,贾玉奎,郝玉光,徐军,高君亮,陈海玲. 干旱胁迫对欧李叶绿素荧光特性的影响[J]. 西北林学院学报,2018,33(6):10−14. Duan N,Jia YK,Hao YG,Xu J,Gao JL,Chen HL. Effects of drought stress on chlorophyll fluorescence characteristics of Cerasus humilis[J]. Journal of Northwest Forestry University,2018,33(6):10−14.
[35] 朱成刚,陈亚宁,李卫红,付爱红,杨玉海. 干旱胁迫对胡杨PSⅡ光化学效率和激能耗散的影响[J]. 植物学报,2011,46(4):413−424. doi: 10.3724/SP.J.1259.2011.00413 Zhu CG,Chen YN,Li WH,Fu AH,Yang YH. Effect of drought stress on photochemical efficiency and dissipation of excited energy in photosystem Ⅱ of Populus euphratica[J]. Chinese Bulletin of Botany,2011,46(4):413−424. doi: 10.3724/SP.J.1259.2011.00413
[36] Liu LT,Li AC,Chen J,Wang M,Zhang YJ,et al. iTRAQ-based quantitative proteomic analysis of cotton (Gossypium hirsutum L.) leaves reveals pathways associated throughout the aging process[J]. Acta Physiol Plant,2019,41(8):144. doi: 10.1007/s11738-019-2921-y
[37] 董高峰,陈贻竹,李耿光,黄涛,阳成伟. 阳生植物和阴生植物的叶黄素循环与非辐射能量耗散[J]. 武汉植物学研究,2001,19(2):128−134. Dong GF,Chen YZ,Li GG,Huang T,Yang CW. Xanthophyll cycle and non-radiative energy dissipation in sun and shade plants[J]. Journal of Wuhan Botanical Research,2001,19(2):128−134.
[38] Lawson T,Blatt MR. Stomatal size,speed,and responsiveness impact on photosynthesis and water use efficiency[J]. Plant Physiol,2014,164(4):1556−1570. doi: 10.1104/pp.114.237107
[39] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报,1999,16(4):444−448. Zhang SR. A discussion on chlorophyll fluorescence kinetics parameters and their significance[J]. Chinese Bulletin of Botany,1999,16(4):444−448.
[40] 王海珍,韩路,徐雅丽,刘艳萍,王琳. 胡杨异形叶光合系统Ⅱ叶绿素荧光特性[J]. 西北植物学报,2019,39(10):1795−1804. doi: 10.7606/j.issn.1000-4025.2019.10.1795 Wang HZ,Han L,Xu YL,Liu YP,Wang L. Chlorophyll fluorescence characteristics of photosystem Ⅱ of Populus euphratica heteromorphic leaves[J]. Acta Botanica Boreali-Occidentalia Sinica,2019,39(10):1795−1804. doi: 10.7606/j.issn.1000-4025.2019.10.1795
[41] 张瑞香. 不同林龄野生南方红豆杉碳氮磷生态化学计量与非结构性碳水化合物特征研究[D]. 郑州:河南农业大学,2023:1−52. [42] Quevedo-Rojas A,García-Núñez C,Jerez-Rico M,Jaimez R,Schwarzkopf T. Leaf acclimation strategies to contrasting light conditions in saplings of different shade tolerance in a tropical cloud forest[J]. Funct Plant Biol,2018,45(9):968−982. doi: 10.1071/FP17308
[43] 孙晓伟,王兴昌,孙慧珍,全先奎,杨青杰. 雌雄异株树种山杨、水曲柳和东北红豆杉光合特性对比[J]. 南京林业大学学报(自然科学版),2023,47(1):129−135. Sun XW,Wang XC,Sun HZ,Quan XK,Yang QJ. Photosynthetic characteristics of dioecious Populus davidiana,Fraxinus mandshurica and Taxus cuspidata[J]. Journal of Nanjing Forestry University (Natural Sciences Edition),2023,47(1):129−135.
-
期刊类型引用(11)
1. 王世彤,宋帅帅,李杰华,杨腾,何艺琴,魏新增,江明喜. 极小种群野生植物黄梅秤锤树的光合生理特性. 生态学杂志. 2024(03): 701-708 .  百度学术
百度学术
2. 魏新增,蒲云海,史红文,肖之强,江明喜. 湖北省国家重点保护野生植物分布与研究进展. 广西植物. 2024(11): 2000-2009 . 百度学术
3. 常二梅,刘建锋,黄跃宁,李红丽,单冰燕,江泽平,赵秀莲. 岷江柏野生居群和迁地保护居群的遗传多样性比较. 植物研究. 2022(05): 772-779 . 百度学术
4. 许玥,臧润国. 中国极小种群野生植物保护理论与实践研究进展. 生物多样性. 2022(10): 84-105 . 百度学术
5. 江全,郑旭,张康,展阳,唐罗忠. 秤锤树的种实基本性状与种子休眠机理研究. 西南林业大学学报(自然科学). 2021(02): 145-150 . 百度学术
6. 谢春平,刘大伟,吴显坤,薛晓明,南程慧. 基于灰色关联度分析的浙江楠在江苏的适宜引种地评估. 云南农业大学学报(自然科学). 2021(02): 330-337 . 百度学术
7. 黄成名,李方俊,苏彩晴,张海玲,李林. 湖北省秤锤树保护性引种试验. 甘肃林业科技. 2020(02): 5-6+27 . 百度学术
8. 杨腾,王世彤,魏新增,江明喜. 中国特有属秤锤树属植物的潜在分布区预测. 植物科学学报. 2020(05): 627-635 . 本站查看
9. 商侃侃,张希金,宋坤. 上海辰山植物园不同生活型木本植物枝叶大小关系的比较. 植物研究. 2020(05): 641-647 . 百度学术
10. 张丽,杨小波,农寿千,李东海,李苑菱,宋佳昱. 两种不同保护模式下坡垒种群发育特征. 生态学报. 2019(10): 3740-3748 . 百度学术
11. 索南邓登,陈卫东,林鹏程. 青藏高原野生濒危药用植物掌裂兰的生境及濒危因素分析. 广西植物. 2019(09): 1166-1179 . 百度学术
其他类型引用(3)
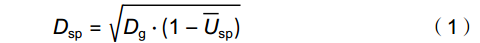





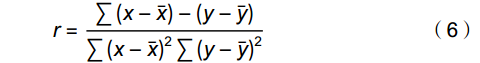

计量
- 文章访问数: 121
- HTML全文浏览量: 20
- PDF下载量: 8
- 被引次数: 14
