
Distribution characteristics and influencing factors of random framework in a northern tropical karst seasonal rainforest
-
摘要:
基于弄岗15-hm2喀斯特季节性雨林动态监测样地2021年的野外复查数据,本研究对林分整体和优势种的随机体分布特征进行分析,并从生活型和环境因子角度探讨随机体的影响因素。结果显示,无论是林分整体还是优势种的角尺度分布中,随机体频率均最高,在0.5以上。林分整体和优势种角尺度分布均为随机分布。乔木分布格局为随机分布,灌木为聚集分布,且随机体比例均超过55%。原有植株个体数、平均胸径、平均海拔、地形湿润度指数和坡度对随机体的解释率较高。综上所述,我们认为,随机体在喀斯特天然林中占据主导地位,其分布受林木生活型、生物因素以及复杂地形条件的共同影响。
Abstract:Based on 2021 field survey data from the 15-hm2 Nonggang karst seasonal rainforest dynamic monitoring plot, this study analyzed the distribution characteristics of random framework within the overall stand and among dominant species. Additionally, the factors influencing the distribution of random framework were explored from the perspectives of life forms and environmental variables. Results showed that the frequency of random framework in the uniform angle index distribution was highest, exceeding 0.5, both for the stand as whole and for dominant species. The overall stand and dominant species both exhibit random distribution. Trees showed a random distribution pattern, while shrubs were aggregated, with random framework accounting for more than 55% of the total. Key factors influencing the occurrence of random framework included the number of original plant individuals, average diameter at breast height, average elevation, topographic wetness index, and slope, which together explained a significant proportion of the variation. In conclusion, random framework occupy a dominant position within karst natural forests, with their distribution significantly influenced by the combined effects of tree life forms, biological factors, and complex topographical conditions.
-
森林空间结构是根据林木的空间位置和属性,来描述林木间相互关系的空间排列方式。而林木个体的空间分布状态,即分布格局,是研究林木在水平空间相互关系的重要指标,同时也是空间结构的基本特征之一[1]。分布格局的研究方法主要有样方法和距离法,但两种方法均存在局限性。
1999年,有学者提出了由参照树和相邻木组成的描述林分空间结构单元的方法[2]。该方法把林区内任意一棵树和距其最近的4棵相邻树组成的结构小组称为林分空间结构单元。角尺度(Wi)就是基于林分空间结构单元所构建的一个参数,用于描述林木的分布格局[3]。角尺度法主要是通过判断由参照树和相邻木构成的夹角大小,来判断相邻木围绕参照树的均匀性。该方法既可以通过图形来判断分布格局,也可以通过数值来判断,其操作方便,计算简单[4]。林木分布格局在林木和森林的生长演替过程中均发挥直接作用。因此,通过分析森林林分整体的角尺度分布特征,能有效地确定林分演替阶段,了解林分的稳定性。同时,对优势种分布格局进行分析,还能了解优势种与其他物种的关系,如竞争、共生、互助等。
随机木是指Wi=0.50的林木,随机木对应的结构单元称为随机体。该概念是Zhang和Hui[5]在2021年基于角尺度参数提出的。随机木在反映林分整体树种组成和竞争情况等方面具有重要作用。研究发现,随机体比例在天然林中不受地域分布、树种组成、林分结构、格局类型和树木竞争的影响,并由此提出了“随机体-稳定性”假说[6]。但在喀斯特区域,因其特有的“富钙、干旱、瘠薄”等异质生境,加之喀斯特复杂的地质地貌和环境因素,使其成为环境异质性与生物物种多样性的独特响应区。因此,生境异质性强的喀斯特地区天然林中,“随机体-稳定性”假说是否仍然成立,还有待检验。Li等[7]分析了喀斯特地区土壤和岩石两种立地上,不同径阶树木群体以及每种立地上两种生活型树木的随机体分布特征,结果发现随机体的比例和分布受生活型和生境异质性的影响,而与树的大小无关。然而,先前对于随机体在喀斯特地区影响因素的分析,均是以尺度较小的样地作为研究对象,在更大的森林样地尺度中,生活型是否对随机体有影响还不清楚。生境异质性的产生主要有两个原因,一是组成生境的能量和物质在时间或空间上的差异,如生物因素、水分状况等[8],二是区域生态系统运动发展的不平衡性[9]。而不同的环境因子在喀斯特地区中对随机体的影响还未知。此外,立地条件对于林木生长也是一个重要因素[10]。因此,研究喀斯特生境中不同因子对随机体的影响差异具有重要意义。
我国桂西南地区是北热带喀斯特季节性雨林的重要分布区之一,不但包含了从山顶到山坡到洼地的各种地形要素,还包含了水平方向上的高异质性以及垂直方向上的多层性,形成了一系列典型的“峰丛-洼地”生境类型[11, 12]。在喀斯特岩溶山地的地形地貌中,存在着复杂多样的小生境,这些小生境受不同因素的影响而呈现出多样性。此外,在不同的地段、坡向和坡位上,小生境的资源分配和植物组成等方面均存在明显差异。本文基于弄岗喀斯特季节性雨林15-hm2样地的第2次复查数据,从林分整体和优势种角尺度分布特征,以及生活型、生物因子和地形因子对随机体的影响进行分析。研究旨在探讨:(1)基于北热带喀斯特季节性雨林相邻木林分整体和优势种的随机体分布特征;(2)不同生活型树种在空间尺度较大的喀斯特生境中对随机体是否有影响;(3)北热带喀斯特季节性雨林不同环境因子对随机体的影响差异。
1. 材料与方法
1.1 研究区概况
弄岗国家级自然保护区地处广西龙州县与宁明县的交界地带,分弄岗、隆呼、陇瑞3个区,总面积为10 077 hm2。保护区土壤类型为原始石灰土、棕色石灰土、黑色石灰土等。年降水集中在5-9月,降水量分布不均匀,干湿交替较强;年平均温度22 ℃,最低温度13 ℃,最高温度37 ℃~39 ℃。
1.2 样地设置与调查
北热带喀斯特季节性雨林地处弄岗自然保护区弄岗片的弄姆皇(22°25′N,106°57′E)。整个样地南北宽300 m,东西长500 m,海拔为180~370 m,平均海拔260 m,样地坡度范围在3.7°~78.9°,平均坡度41.7°。整个样地生境多样化,属于喀斯特“峰丛-洼地”典型的生境类型,在中国森林生物多样性监测网络中,该样地是唯一的典型热带喀斯特森林监测样地。
根据CTFS全球森林生物多样性监测标准,利用全站仪将15-hm2样地划分成1 500个10 m×10 m的样方。在此基础上,于2011年完成了首次植被调查,对样方内胸径(DBH)≥1 cm个体的名称、胸径、坐标及生长状态等进行了调查并记录。之后,每5年进行一次调查,并于2021年完成第2次复查。本文以2021年的复查数据为依据,选取DBH≥5 cm的个体作为研究对象。
1.3 树种优势度的计算
树种优势度(Dsp)指树种在林分中的数量优势度和空间上的优势度,选取优势度位于前10的物种作为优势种,Dsp的计算公式为:
Dsp=√Dg⋅(1−¯Usp) (1) 式中,Dg表示相对显著度,
ˉUsp 表示树种大小比数均值。Dsp接近1表示非常优势,接近0表示几乎没有优势[13]。树种大小比数均值表达式如下:
¯Usp=1Nsp∑Nspi=1Ui (2) 式中,Nsp表示所观察树种的参照树数量,Ui为树种的第i株树的大小比数值。
1.4 角尺度计算
角尺度(Wi)指的是参照树与4株最近邻木间小于标准角的夹角占比,用下式表示[3]:
Wi=1n∑nj=1Zij (3) 其中:Zij=
1{1(当j个角小于标准角α0)0(否则) 式中,Wi为参照树i的角尺度,n指相邻木个体数,n=4。Zij为参照树i与其相邻木j之间的第j个夹角α与标准角α0之间的大小关系,有两种可能的值,当Zij=1时,表明第j个α角小于标准角,当Zij=0时,表明第j个α角大于标准角。
角尺度均值(
¯W )能够反映一个林分整体的分布情况。其计算公式如下:¯W=1N∑Niwi (4) 优势种角尺度均值用下式表示:
¯Wsp=1Nsp∑Nspi=1 (5) 式中,N为样地林木总株数。当角尺度均值在[0.457,0.517]时为随机分布;大于0.517为聚集分布;小于0.475为均匀分布[14]。角尺度等于0.50的林木被称为随机木,相应林木的结构单元称为随机体[15]。角尺度为0.75或1的林木为聚集木,相应林木结构单元称为聚集体。角尺度为0或0.25的林木被称作均匀木,相应林木的结构单元被称作均匀体。本研究在计算林分整体和优势种角尺度时,设置了5 m缓冲区,并将缓冲区内的林木做相邻木处理,以此消除林木的边缘效应造成的影响。
1.5 Pearson 相关分析
运用Pearson相关分析法,判断随机体与环境因子及环境因子间的相关性。相关系数r计算公式如下:
r=∑(x−ˉx)−(y−ˉy)∑(x−ˉx)2∑(y−ˉy)2 (6) r取值范围为−1~1,当|r|≥0.8,表示两者高度相关;0.5≤|r|<0.8,为中度相关;0.3≤|r|<0.5,表示低相关;当|r|<0.3,表示相关性非常微弱,可视为没有相关性,P<0.05表明有统计学意义。
1.6 环境因子获取
用全站仪获取精确海拔数据,计算每个20 m×20 m样方的平均海拔、坡向、坡度和凹凸度4个地形指标[16-18]。土壤湿度状况以地形湿润指数和干旱度指数为表征,参见Punchi-Manage等[19]的研究。样地内岩石裸露情况采用踏查法进行评估。综上共有7个地形因子。此外,还考查了每个样方内最大胸径、胸高断面积之和、植株个体数和平均胸径4个生物因子,共11个环境因子。
1.7 冗余分析
冗余分析的响应变量由每个样方的随机体个数组成。为了消除林木边缘效应的影响,设置2 m缓冲区,缓冲区内的林木做相邻木处理。解释变量数据包括11个环境因子。冗余分析前,对数据进行标准化处理,并比较自变量的相关系数是否大于7,以此衡量自变量间的共线性。再计算每个变量的方差膨胀因子,当方差膨胀因子<10时,表示无明显的共线性问题[20],可进行冗余分析。采用层次分割方法对冗余分析结果进行变差分解,以获取每个解释变量的校正解释率。
1.8 数据处理
数据分析计算均利用R 4.3.1软件完成,冗余分析在vegan程序包中进行,层次分割采用rdacca.hp程序包[21],利用ForestSAS程序包计算角尺度和大小比数均值[22],Pearson相关系数图在corrplot包中绘制,角尺度分布特征图在ggplot2包中完成,单独效应比例图在Origin 2021软件中绘制。
2. 结果与分析
2.1 林分整体和优势种随机体的比例和分布特征
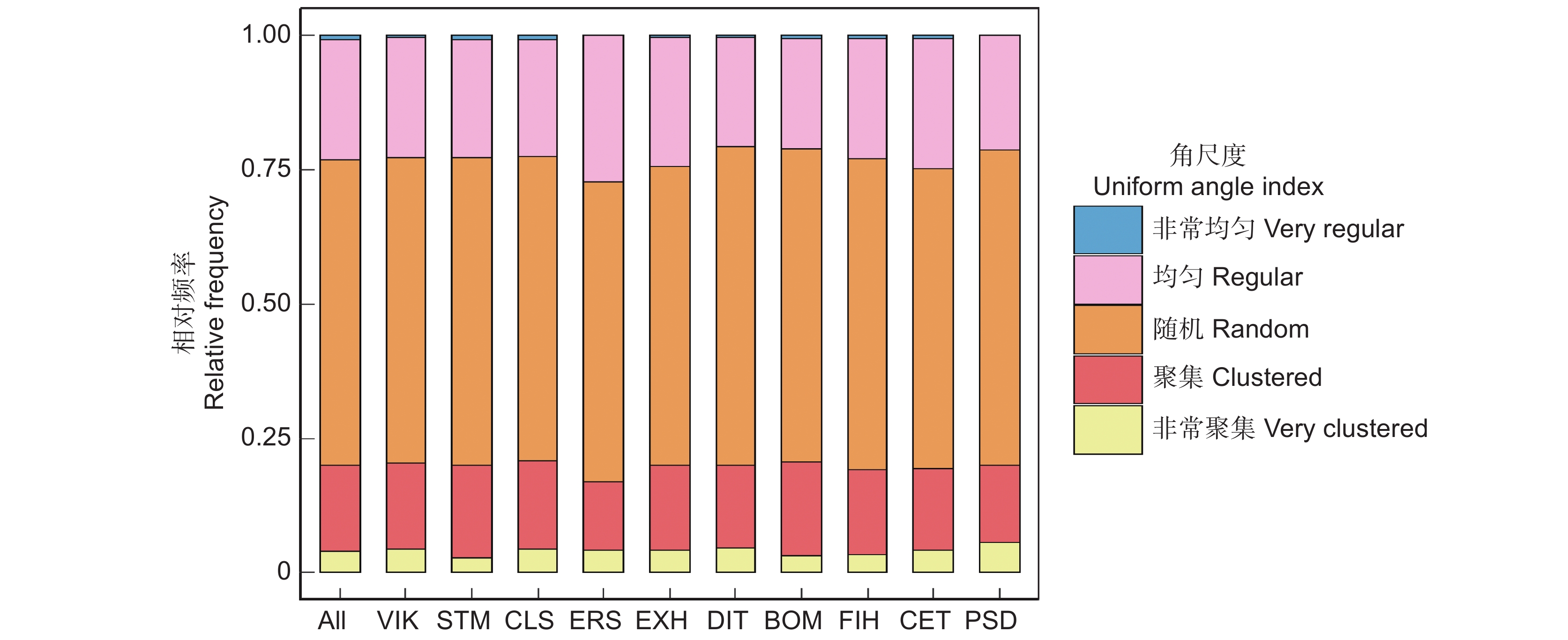
树种优势度位于前10的物种在森林中起关键作用,尤其是闭花木(Cleistanthus sumatranus (Miq.) Muell. Arg.)、苹婆(Sterculia monosperma Ventenat)和广西牡荆(Vitex kwangsiensis P'ei),不仅在数量上占林分整体的绝大多数,其优势度也是最高的,分别为0.262、0.260、0.245,其他树种的优势度均小于0.200。角尺度计算结果表明,林分整体和优势种角尺度均值都在随机分布范围内(表1)。但林分整体和优势种随机木的分布频率均超过0.5,而均匀木和聚集木分布频率均比随机木低,说明样地中大多数林木的最近相邻木处于随机分布,各个优势种的最近相邻木处于随机分布的也占大多数,即随机体占绝对分布(图1)。
表 1 北热带喀斯特季节性雨林树种优势度排名前10的树种Table 1. Top 10 dominant species in northern tropical karst seasonal rainforest树种
Species科名
Family株数
Number of
plants大小比数
Dominance相对显著度
Relative
significance优势度
Advantage
degree角尺度
Uniform angle
index广西牡荆
Vitex kwangsiensis P'ei唇形科 1 645 0.325 0.122 0.286 0.504 苹婆
Sterculia monosperma Ventenat梧桐科 2 426 0.491 0.134 0.262 0.498 闭花木
Cleistanthus sumatranus (Miq.) Muell. Arg.大戟科 3 054 0.530 0.139 0.256 0.504 劲直刺桐
Erythrina stricta Roxb.豆科 242 0.074 0.031 0.171 0.485 蚬木
Excentrodendron tonkinense (A. Chev.)
H. T. Chang et R. H. Miau锦葵科 483 0.367 0.039 0.156 0.498 海南椴
Diplodiscus trichosperma (Merrill) Y. Tang锦葵科 456 0.317 0.033 0.150 0.509 黄梨木
Boniodendron minus (Hemsl.) T. Chen无患子科 404 0.314 0.026 0.133 0.506 对叶榕
Ficus hispida L. f.桑科 676 0.602 0.031 0.110 0.497 假玉桂
Celtis timorensis Span大麻科 414 0.509 0.020 0.099 0.495 鱼骨木
Psydrax dicocca Gaertn.茜草科 252 0.405 0.015 0.094 0.510 林分整体 − 10 052 0.489 − − 0.500 ![]() 图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.
图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforestAll: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.2.2 不同生活型树种随机体的比例和分布特征
乔木角尺度均值在随机分布范围内,为0.499,随机体占比为57.167%,均匀体占比较小,为23.189%,聚集体次之,为19.644%。灌木角尺度均值为0.519,为聚集分布,随机体占比为59.173%,均匀体占比为19.064%,聚集体为21.763%。乔木和灌木的随机体比例均超过55%,但灌木的聚集体占比高于均匀体。而在乔木中,均匀体占比高于聚集体,表明不同生活型的植物,其随机体比例和分布存在一定的差异(表2)。
表 2 北热带喀斯特季节性雨林不同生活型随机体比例和分布Table 2. Proportion and distribution of random framework among different life forms in northern tropical karst seasonal rainforest生活型
Life style角尺度 Uniform angle index / % 非常均匀
Very regular均匀
Regular随机
Random聚集
Clustered非常聚集
Very clustered角尺度均值
Uniform angle index mean乔木 0.773 22.416 57.167 15.785 3.859 0.499 灌木 0.719 18.345 59.173 16.187 5.576 0.519 2.3 Pearson相关性分析
研究结果显示,随机体与胸高断面积之和、平均海拔、坡度、岩石裸露率、凹凸度、干旱度指数、原有植株个体数之间均存在显著正相关,其中相关性最高的为原有植株个体数,相关系数达0.86;平均海拔和坡度分别与随机体存在低度相关,相关系数分别为0.36和0.31。随机体分别与最大胸径、平均胸径和地形湿润指数存在显著负相关,但与坡向无显著相关性。胸高断面积和最大胸径、坡度和地形湿润指数间也具有较高的相关性,相关系数分别为0.7和−0.72(图2)。
![]() 图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.
图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforestMeanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.2.4 环境因子与随机体的冗余分析及单个解释变量的相对重要性
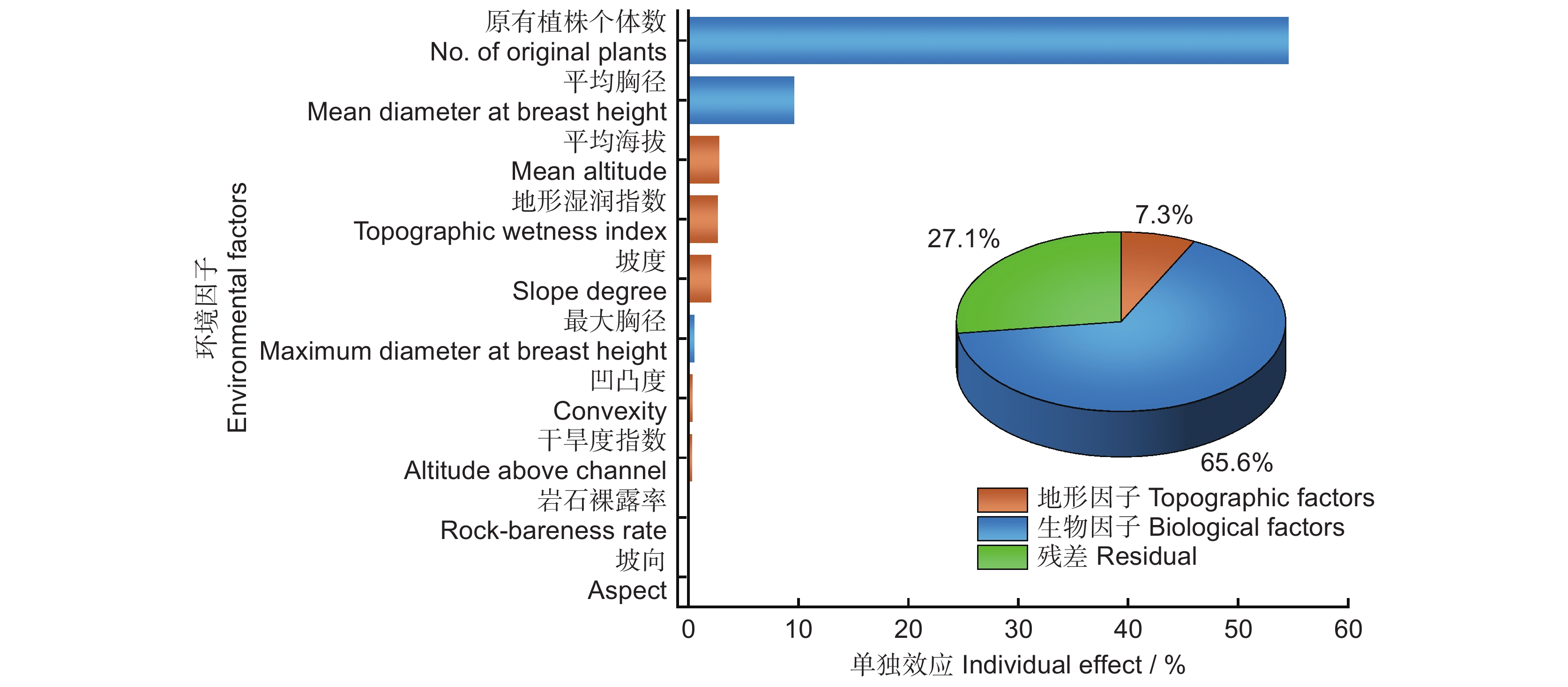
结合环境因子的相关性和方差膨胀因子分析,将胸高断面积之和剔除。对随机体与生物因子和地形因子进行冗余分析,10个环境因子对随机体的解释量R2经校正后为0.729。其中,生物因子解释了65.6%,地形因子解释了7.3%。未被解释的部分占27.1%。根据层次分割法各环境因子单独效应比例排序,随机体的主要影响因素包括原有植株个体数、平均胸径、平均海拔、地形湿润指数和坡度;贡献较小的是最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数(图3)。
![]() 图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest
图 3 北热带喀斯特季节性雨林环境因子单独效应比例图Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest3. 讨论
3.1 随机体的分布特征
林木的空间分布格局主要分为随机分布、规则分布和聚集分布,最理想的是随机分布,而随机分布也是天然原始顶极群落中最为显著的结构特征[23]。本研究中,林分整体为随机分布,优势种的角尺度均值也在随机分布范围内,且随机体占比均超过50%。因此,随机体以相似的比例出现在不同树种中,且支持了“随机体-稳定性”假说,与前人研究结果一致[5]。这种结构体相对来说具有较小的生存压力,稳定性高,在竞争情况下不易出现弱势或者不健康的林木,在自然演替过程中存活几率更大[6]。而均匀体和聚集体占比较小,与袁星明等[24]的报道相似。
不同空间分布格局分析方法可能导致结果上的差异。前人研究采用完全随机零模型和泊松异质性零模型对该地区分布格局进行了分析,完全随机零模型结果表明,在小尺度上所有物种均呈聚集分布[25],与本研究结果不一致;然而,当泊松异质性零模型去除生境异质性后,大部分物种表现为随机分布,与本文结果一致。完全随机零模型是基于假设个体随机分布,不受任何因素的干扰而对分布格局进行拟合分析。但在生境异质性强的北热带喀斯特地区,物种的分布显然会受到各种因素的影响。因此,在不考虑任何因素干扰情况下大多数物种呈聚集分布,与本研究结果存在差异。泊松异质性零模型主要用于模拟生境关联,本研究基于相邻木的角尺度法分析林木分布格局,也未直接考虑生境异质性,所以二者结果一致。另外,角尺度法不存在尺度效应,而空间点格局法可以拟合分析不同尺度的林木分布格局,二者各具特点。对于这两种方法,有学者也进行了对比,发现角尺度法在判断分布格局的准确性、有效性和可行性方面优于空间点格局分析[26]。
林分密度是预测林分动态的重要空间信息之一[27]。以往研究表明,种群分布格局对密度具有一定的依赖性,密度越大,聚集强度越高[28]。在本研究中,闭花木的密度最大,聚集程度也是最高的。尽管苹婆与鱼骨木在数量上存在显著差异,但它们在聚集体中所占的比例却几乎相同。这可能是在北热带喀斯特季节性雨林中岩石的裸露率较高,在岩石裸露度高的区域,植物聚集度就会偏低,反之亦然。因此,种群的分布格局与密度无明显的线性关系。
3.2 生活型对随机体的影响
植物生活型的形成是植物在特定环境中趋同适应的结果。灌木主要处于上层乔木树种的林冠下或林隙中,在数量特征和空间分布格局上会受后者的制约与影响[29]。本研究中,乔木分布比灌木多,优势度位于前10的皆为乔木树种,生活型为灌木的树种仅占25.8%。灌木树种相对于乔木树种其聚集体比例更高,分布格局为聚集分布,而乔木树种为随机分布。研究表明,不同生活型的种群聚集度差异很大,从乔木层到灌木层,聚集度逐渐增加,在林下灌丛层中,聚集度最高[25],与本文结果一致。因此随机体分布与树种自身的生物学特性有一定关系。
3.3 环境因子对随机体的影响
随机体受诸多因素的影响,不同因素的影响程度不同。本研究中,植株个体数、平均胸径、平均海拔、地形湿润指数和坡度对随机体的影响较大,而最大胸径、坡向、岩石裸露率、凹凸度和干旱度指数对随机体的影响较小。从相关系数可以看出,植株个体数与随机体呈显著正相关,植株的个体数越多,随机体所占比例越高,群落越稳定。
林木间的竞争是最直观的相互影响关系。在林区,因为森林资源有限,为了生存,林木之间必然要进行争夺[30-32]。林木自身遗传物质的差异,导致了林木自身的竞争能力也不一样。竞争力可以从林木的胸径等属性中反映出来[33]。不同树种之间的竞争强度会影响其生长状况,从而影响其在森林中的分布。此外,强烈的竞争还会造成一些树木的死亡,从而使森林随机体的分布发生改变。本研究中优势种随机体的分布情况不一,既存在随机分布,也存在聚集分布。因此,竞争是影响随机体分布的重要原因之一。
海拔对植物的分布起重要作用,一方面是直接作用,另一方面是对其他非生物资源(如光、温、水、土等)的分配产生影响。然而,在本研究中,地形因子对于随机体分布的解释力度相对较低。尽管如此,在这些地形因子之中,海拔仍然是解释随机体分布最具影响力的因素。在北热带喀斯特季节性雨林中,物种丰富度随着海拔梯度的变化而呈现显著差异[34],物种之间的竞争强度和自疏程度都有所提高,而同种植物之间的聚集程度则有所下降[35]。同时,由于海拔的不同,水热条件也会发生较大的差异,从而使植物在不同海拔有明显的分布差异。在海拔较高的地区,如山顶,其光照强度高、水分缺乏、昼夜温差大、岩石裸露度高、土层薄。而在洼地边缘通常伴有季节性水淹,光照强度较弱[36]。因此,海拔直接或间接影响林木分布[37],造成随机体分布的差异。
4. 结论
本研究分析了北热带喀斯特季节性雨林森林动态监测样地中的随机体分布特征及其影响因素,发现该区林分整体稳定性较高,多数林木处于随机分布状态。其中,闭花木、苹婆和广西牡荆树种占主要优势,且其角尺度对林分整体分布起关键作用。不同生活型植物的随机体分布不同,树种本身生物学特性是影响不同生活型树种随机体分布的原因。不同环境因子对随机体的影响程度不同,原有植株个体数、平均胸径、平均海拔、地形湿润度指数和坡度是随机体的主要驱动因子。
-
![]()
图 1 北热带喀斯特季节性雨林林分整体和优势种角尺度分布特征
All :林分整体 ;VIK :广西牡荆 ;STM :苹婆;CLS: 闭花木 ; ERS :劲直刺桐; EXH :蚬木 ; DIT :海南椴 ;BOM :黄 梨木 ; FIH :对叶榕 ;CET :假玉桂 ;PSD :鱼骨木。
Figure 1. Uniform angle index distribution characteristics of overall stand and dominant species in northern tropical karst seasonal rainforest
All: Entire stand; VIK: Vitex kwangsiensis C. P'ei; STM: Sterculia monosperma Vent; CLS: Cleistanthus sumatranus (Miq.) Müll. Arg.; ERS: Erythrina stricta Roxb.; EXH: Excentrodendron hsienmu (A. Chev.) H. T. Chang et R. H. Miao; DIT: Diplodiscus trichospermus (Merr.) Y. Tang, M. G. Gilbert et Dorr; BOM: Boniodendron minus (Hemsl.) T.C. Chen; FIH: Ficus hispida L. f.; CET: Celtis timorensis Span.; PSD: Psydrax dicocca Gaertn.
![]()
图 2 北热带喀斯特季节性雨林随机体与环境因子Pearson相关性矩阵图
Meanelev:平均海拔;Slope:坡度;Aspect:坡向;Convex:凹凸度;TWI:地形湿润指数;ACH:干旱度指数;RBR:岩石裸露率;ba:胸高断面积之和;Maxdbh:最大胸径;Meandbh:平均胸径。
Figure 2. Pearson correlation matrix between random framework and environmental factors in northern tropical karst seasonal rainforest
Meanelev: Mean elevation; Slope: Slope; Aspect: Aspect; Convex: Convexity; TWI: Topographic wetness index; ACH: Altitude above channel; RBR: Rock-bareness rate; ba: Total basal area; Maxdbh: Maximum diameter at breast height; Meandbh: Mean diameter at breast height.
![]()
图 3 北热带喀斯特季节性雨林环境因子单独效应比例图
Figure 3. Individual effect proportions of environmental factors in northern tropical karst seasonal rainforest
表 1 北热带喀斯特季节性雨林树种优势度排名前10的树种
Table 1 Top 10 dominant species in northern tropical karst seasonal rainforest
树种
Species科名
Family株数
Number of
plants大小比数
Dominance相对显著度
Relative
significance优势度
Advantage
degree角尺度
Uniform angle
index广西牡荆
Vitex kwangsiensis P'ei唇形科 1 645 0.325 0.122 0.286 0.504 苹婆
Sterculia monosperma Ventenat梧桐科 2 426 0.491 0.134 0.262 0.498 闭花木
Cleistanthus sumatranus (Miq.) Muell. Arg.大戟科 3 054 0.530 0.139 0.256 0.504 劲直刺桐
Erythrina stricta Roxb.豆科 242 0.074 0.031 0.171 0.485 蚬木
Excentrodendron tonkinense (A. Chev.)
H. T. Chang et R. H. Miau锦葵科 483 0.367 0.039 0.156 0.498 海南椴
Diplodiscus trichosperma (Merrill) Y. Tang锦葵科 456 0.317 0.033 0.150 0.509 黄梨木
Boniodendron minus (Hemsl.) T. Chen无患子科 404 0.314 0.026 0.133 0.506 对叶榕
Ficus hispida L. f.桑科 676 0.602 0.031 0.110 0.497 假玉桂
Celtis timorensis Span大麻科 414 0.509 0.020 0.099 0.495 鱼骨木
Psydrax dicocca Gaertn.茜草科 252 0.405 0.015 0.094 0.510 林分整体 − 10 052 0.489 − − 0.500  下载: 导出CSV
下载: 导出CSV
表 2 北热带喀斯特季节性雨林不同生活型随机体比例和分布
Table 2 Proportion and distribution of random framework among different life forms in northern tropical karst seasonal rainforest
生活型
Life style角尺度 Uniform angle index / % 非常均匀
Very regular均匀
Regular随机
Random聚集
Clustered非常聚集
Very clustered角尺度均值
Uniform angle index mean乔木 0.773 22.416 57.167 15.785 3.859 0.499 灌木 0.719 18.345 59.173 16.187 5.576 0.519
下载: 导出CSV
-
[1] 刘帅,李建军,李丹,朱凯文,郭瑞,等. 林木空间分布格局分析方法比较及其适应性[J]. 林业科学,2019,55(11):73−84. doi: 10.11707/j.1001-7488.20191109 Liu S,Li JJ,Li D,Zhu KW,Guo R,et al. Comparison and adaptability of analytical methods for spatial distribution patterns in forest[J]. Scientia Silvae Sinicae,2019,55(11):73−84. doi: 10.11707/j.1001-7488.20191109
[2] 惠刚盈. 基于相邻木关系的林分空间结构参数应用研究[J]. 北京林业大学学报,2013,35(4):1−9. Hui GY. Studies on the application of stand spatial structure parameters based on the relationship of neighborhood trees[J]. Journal of Beijing Forestry University,2013,35(4):1−9.
[3] 惠刚盈. 角尺度 —— 一个描述林木个体分布格局的结构参数[J]. 林业科学,1999,35(1):37−42. Hui GY. The neighbourhood pattern-a new structure parameter for describing distribution of forest tree position[J]. Scientia Silvae Sinicae,1999,35(1):37−42.
[4] Pommerening A. Evaluating structural indices by reversing forest structural analysis[J]. Forest Ecol Manag,2006,224(3):266−277. doi: 10.1016/j.foreco.2005.12.039
[5] Zhang GQ,Hui GY. Random trees are the cornerstones of natural forests[J]. Forests,2021,12(8):1046. doi: 10.3390/f12081046
[6] 惠刚盈,赵中华,张弓乔,胡艳波. 基于角尺度的随机体在森林稳定性维持中的作用[J]. 林业科学,2021,57(2):22−30. doi: 10.11707/j.1001-7488.20210203 Hui GY,Zhao ZH,Zhang GQ,Hu YB. The role of random structural pattern based on uniform angle index in maintaining forest stability[J]. Scientia Silvae Sinicae,2021,57(2):22−30. doi: 10.11707/j.1001-7488.20210203
[7] Li YF,Luo XQ,Li J. Habitat heterogeneity in karst environments influences the proportion and distribution of random framework[J]. Ecol Ind,2022,143:109387. doi: 10.1016/j.ecolind.2022.109387
[8] Ben-Hur E,Kadmon R. Heterogeneity–diversity relationships in sessile organisms:a unified framework[J]. Ecol Lett,2020,23(1):193−207. doi: 10.1111/ele.13418
[9] 龙健,吴求生,李娟,廖洪凯,刘灵飞,等. 贵州茂兰喀斯特森林不同小生境类型对岩石溶蚀的影响[J]. 土壤学报,2021,58(1):151−161. Long J,Wu QS,Li J,Liao HK,Liu LF,et al. Effects of different microhabitat types on rock dissolution in Maolan Karst Forest,Southwest China[J]. Acta Pedologica Sinica,2021,58(1):151−161.
[10] 范叶青,周国模,施拥军,杜华强,周宇峰,徐小军. 地形条件对毛竹林分结构和植被碳储量的影响[J]. 林业科学,2013,49(11):177−182. Fan YQ,Zhou GM,Shi YJ,Du HQ,Zhou YF,Xu XJ. Effects of terrain on stand structure and vegetation carbon storage of phyllostachys edulis forest[J]. Scientia Silvae Sinicae,2013,49(11):177−182.
[11] 王斌,黄俞淞,李先琨,向悟生,丁涛,等. 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布[J]. 生物多样性,2014,22(2):141−156. doi: 10.3724/SP.J.1003.2014.13195 Wang B,Huang YS,Li XK,Xiang WS,Ding T,et al. Species composition and spatial distribution of a 15 ha northern tropical karst seasonal rain forest dynamics study plot in Nonggang,Guangxi,southern China[J]. Biodiversity Science,2014,22(2):141−156. doi: 10.3724/SP.J.1003.2014.13195
[12] 郭屹立,王斌,向悟生,丁涛,陆树华,等. 弄岗喀斯特季节性雨林15 ha样地密度制约效应分析[J]. 科学通报,2015,60(17):1602−1611. doi: 10.1360/N972014-00892 Guo YL,Wang B,Xiang WS,Ding T,Lu SH,et al. Density-dependent effects of tree species in a 15 ha seasonal rain forest plot in northern tropical karst in Nonggang,Guangxi,southern China[J]. Chinese Science Bulletin,2015,60(17):1602−1611. doi: 10.1360/N972014-00892
[13] 惠刚盈,胡艳波,刘瑞红. 森林观察研究中的林分空间优势度分析方法[J]. 温带林业研究,2019,2(1):1−6,12. Hui GY,Hu YB,Liu RH. Methods of analyzing stand spatial dominance in forest observational studies[J]. Journal of Temperate Forestry Research,2019,2(1):1−6,12.
[14] Hui GY,von Gadow K. Das Winkelmaß Herleitung des optimalen Standardwinkels[J]. Allg Forst Jagdztg,2002,173(10):173−177.
[15] Zhang GQ,Hui GY,Zhao ZH,Hu YB,Wang HX,et al. Composition of basal area in natural forests based on the uniform angle index[J]. Ecol Inform,2018,45:1−8. doi: 10.1016/j.ecoinf.2018.01.002
[16] Harms KE,Condit R,Hubbell SP,Foster RB. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot[J]. J Ecol,2001,89(6):947−959. doi: 10.1111/j.1365-2745.2001.00615.x
[17] Valencia R,Foster RB,Villa G,Condit R,Svenning JC,et al. Tree species distributions and local habitat variation in the Amazon:large forest plot in eastern Ecuador[J]. J Ecol,2004,92(2):214−229. doi: 10.1111/j.0022-0477.2004.00876.x
[18] 郭屹立,王斌,向悟生,丁涛,陆树华,等. 桂西南喀斯特季节性雨林枯立木的空间格局及生境关联性分析[J]. 广西植物,2016,36(2):154−161. doi: 10.11931/guihaia.gxzw201507024 Guo YL,Wang B,Xiang WS,Ding T,Lu SH,et al. Spatial distribution and habitat-association of snags in the tropical karst seasonal rainforest of Southwest Guangxi,China[J]. Guihaia,2016,36(2):154−161. doi: 10.11931/guihaia.gxzw201507024
[19] Punchi-Manage R,Getzin S,Wiegand T,Kanagaraj R,Gunatilleke CVS,et al. Effects of topography on structuring local species assemblages in a Sri Lankan mixed dipterocarp forest[J]. J Ecol,2013,101(1):149−160. doi: 10.1111/1365-2745.12017
[20] 邓婷婷,魏岩,任思远,祝燕. 北京东灵山暖温带落叶阔叶林地形和林分结构对林下草本植物物种多样性的影响[J]. 生物多样性,2023,31(7):22671. doi: 10.17520/biods.2022671 Deng TT,Wei Y,Ren SY,Zhu Y. Effects of topography and stand structure of warm temperate deciduous broad-leaved forest on understory herb diversity in Donglingshan,Beijing[J]. Biodiversity Science,2023,31(7):22671. doi: 10.17520/biods.2022671
[21] 刘尧,于馨,于洋,胡文浩,赖江山. R程序包“rdacca. hp”在生态学数据分析中的应用:案例与进展[J]. 植物生态学报,2023,47(1):134−144. doi: 10.17521/cjpe.2022.0314 Liu Y,Yu X,Yu Y,Hu WH,Lai JS. Application of “rdacca. hp” R package in ecological data analysis:case and progress[J]. Chinese Journal of Plant Ecology,2023,47(1):134−144. doi: 10.17521/cjpe.2022.0314
[22] 柴宗政. 基于相邻木关系的森林空间结构量化评价及R语言编程实现——以秦岭中山带典型次生林为例[D]. 杨凌:西北农林科技大学,2016:1−30. [23] 惠刚盈,赵中华,胡艳波,张弓乔,程世平,徐雪飞. 基于结构参数均值的林分空间结构综合评价研究[J]. 林业科学研究,2023,36(2):12−21. Hui GY,Zhao ZH,Hu YB,Zhang GQ,Cheng SP,Xu XF. Comprehensive evaluation of forest spatial structure based on the mean values of structural parameters[J]. Forest Research,2023,36(2):12−21.
[24] 袁星明,朱宁华,周光益,党鹏,尚海. 湘西喀斯特地区42年生湿地松-樟树人工混交林空间结构研究[J]. 中南林业科技大学学报,2022,42(4):49−58. Yuan XM,Zhu NH,Zhou GY,Dang P,Shang H. Spatial structure of the 42-year-old Pinus elliottii-Cinnamomum camphora mixed plantation in the karst area of western Hunan[J]. Journal of Central South University of Forestry & Technology,2022,42(4):49−58.
[25] 郭屹立,王斌,向悟生,丁涛,陆树华,等. 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析[J]. 生物多样性,2015,23(2):183−191. doi: 10.17520/biods.2014126 Guo YL,Wang B,Xiang WS,Ding T,Lu SH,et al. Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang,Guangxi,southern China[J]. Biodiversity Science,2015,23(2):183−191. doi: 10.17520/biods.2014126
[26] 惠刚盈,李丽,赵中华,党普兴. 林木空间分布格局分析方法[J]. 生态学报,2007,27(11):4717−4728. doi: 10.3321/j.issn:1000-0933.2007.11.040 Hui GY,Li L,Zhao ZH,Dang PX. The comparison of methods in analysis of the tree spatial distribution pattern[J]. Acta Ecologica Sinica,2007,27(11):4717−4728. doi: 10.3321/j.issn:1000-0933.2007.11.040
[27] Pretzsch H,Biber P. Tree species mixing can increase maximum stand density[J]. Can J For Res,2016,46(10):1179−1193. doi: 10.1139/cjfr-2015-0413
[28] Chen P,Xia JB,Ma HS,Gao FL,Dong MM,et al. Analysis of spatial distribution pattern and its influencing factors of the Tamarix chinensis population on the beach of the muddy coastal zone of Bohai Bay[J]. Ecol Ind,2022,140:109016. doi: 10.1016/j.ecolind.2022.109016
[29] Piao T,Comita LS,Jin GZ,Kim JH. Density dependence across multiple life stages in a temperate old-growth forest of northeast China[J]. Oecologia,2013,172(1):207−217. doi: 10.1007/s00442-012-2481-y
[30] Biging GS,Dobbertin M. A comparison of distance-dependent competition measures for height and basal area growth of individual conifer trees[J]. Forest Sci,1992,38(3):695−720. doi: 10.1093/forestscience/38.3.695
[31] Tomé M,Burkhart HE. Distance-dependent competition measures for predicting growth of individual trees[J]. Forest Sci,1989,35(3):816−831. doi: 10.1093/forestscience/35.3.816
[32] Contreras MA,Affleck D,Chung W. Evaluating tree competition indices as predictors of basal area increment in western Montana forests[J]. Forest Ecol Manag,2011,262(11):1939−1949. doi: 10.1016/j.foreco.2011.08.031
[33] Opio C,Jacob N,Coopersmith D. Height to diameter ratio as a competition index for young conifer plantations in northern British Columbia,Canada[J]. Forest Ecol Manag,2000,137(1-3):245−252. doi: 10.1016/S0378-1127(99)00312-6
[34] 黄甫昭,丁涛,李先琨,郭屹立,王斌,等. 弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化[J]. 生态学报,2016,36(14):4509−4517. Huang FZ,Ding T,Li XK,Gou YL,Wang B,et al. Species diversity for various associations along an altitudinal gradient in the karst seasonal rainforest in Nonggang[J]. Acta Ecologica Sinica,2016,36(14):4509−4517.
[35] 龙俊松,汤孟平. 天目山常绿阔叶林空间结构与地形因子的关系[J]. 浙江农林大学学报,2021,38(1):47−57. doi: 10.11833/j.issn.2095-0756.20200267 Long JS,Tang MP. Relationship between spatial structure and terrain factors of evergreen broad-leaved forest in Mount Tianmu[J]. Journal of Zhejiang A&F University,2021,38(1):47−57. doi: 10.11833/j.issn.2095-0756.20200267
[36] Guo YL,Wang B,Mallik AU,Huang FZ,Xiang WS,et al. Topographic species–habitat associations of tree species in a heterogeneous tropical karst seasonal rain forest,China[J]. J Plant Ecol,2017,10(3):450−460.
[37] 赵雪,刘妍妍,金光泽. 地形对阔叶红松林幼苗更新的影响[J]. 应用生态学报,2013,24(11):3035−3042. Zhao X,Liu YY,Jin GZ. Effects of topography on seedling regeneration in a mixed broadleaved-Korean pine forest in Xiaoxing’ an Mountains,Northeast China[J]. Chinese Journal of Applied Ecology,2013,24(11):3035−3042.
-
期刊类型引用(11)
1. 王世彤,宋帅帅,李杰华,杨腾,何艺琴,魏新增,江明喜. 极小种群野生植物黄梅秤锤树的光合生理特性. 生态学杂志. 2024(03): 701-708 .  百度学术
百度学术
2. 魏新增,蒲云海,史红文,肖之强,江明喜. 湖北省国家重点保护野生植物分布与研究进展. 广西植物. 2024(11): 2000-2009 . 百度学术
3. 常二梅,刘建锋,黄跃宁,李红丽,单冰燕,江泽平,赵秀莲. 岷江柏野生居群和迁地保护居群的遗传多样性比较. 植物研究. 2022(05): 772-779 . 百度学术
4. 许玥,臧润国. 中国极小种群野生植物保护理论与实践研究进展. 生物多样性. 2022(10): 84-105 . 百度学术
5. 江全,郑旭,张康,展阳,唐罗忠. 秤锤树的种实基本性状与种子休眠机理研究. 西南林业大学学报(自然科学). 2021(02): 145-150 . 百度学术
6. 谢春平,刘大伟,吴显坤,薛晓明,南程慧. 基于灰色关联度分析的浙江楠在江苏的适宜引种地评估. 云南农业大学学报(自然科学). 2021(02): 330-337 . 百度学术
7. 黄成名,李方俊,苏彩晴,张海玲,李林. 湖北省秤锤树保护性引种试验. 甘肃林业科技. 2020(02): 5-6+27 . 百度学术
8. 杨腾,王世彤,魏新增,江明喜. 中国特有属秤锤树属植物的潜在分布区预测. 植物科学学报. 2020(05): 627-635 . 本站查看
9. 商侃侃,张希金,宋坤. 上海辰山植物园不同生活型木本植物枝叶大小关系的比较. 植物研究. 2020(05): 641-647 . 百度学术
10. 张丽,杨小波,农寿千,李东海,李苑菱,宋佳昱. 两种不同保护模式下坡垒种群发育特征. 生态学报. 2019(10): 3740-3748 . 百度学术
11. 索南邓登,陈卫东,林鹏程. 青藏高原野生濒危药用植物掌裂兰的生境及濒危因素分析. 广西植物. 2019(09): 1166-1179 . 百度学术
其他类型引用(3)
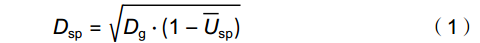





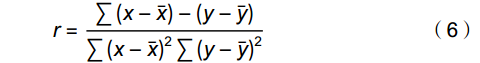

计量
- 文章访问数: 174
- HTML全文浏览量: 26
- PDF下载量: 35
- 被引次数: 14
