Zhou SN,Li JK,Tang LY,Li MX,Cui YX,He GZ,Su J. Effects of mineral elements on the growth and development of Amomum villosum Lour. plants[J]. Plant Science Journal,2023,41(1):89−101. DOI: 10.11913/PSJ.2095-0837.22080
Citation:
Zhou SN,Li JK,Tang LY,Li MX,Cui YX,He GZ,Su J. Effects of mineral elements on the growth and development of Amomum villosum Lour. plants[J]. Plant Science Journal,2023,41(1):89−101. DOI: 10.11913/PSJ.2095-0837.22080
Zhou SN,Li JK,Tang LY,Li MX,Cui YX,He GZ,Su J. Effects of mineral elements on the growth and development of Amomum villosum Lour. plants[J]. Plant Science Journal,2023,41(1):89−101. DOI: 10.11913/PSJ.2095-0837.22080
Citation:
Zhou SN,Li JK,Tang LY,Li MX,Cui YX,He GZ,Su J. Effects of mineral elements on the growth and development of Amomum villosum Lour. plants[J]. Plant Science Journal,2023,41(1):89−101. DOI: 10.11913/PSJ.2095-0837.22080
College of Chinese Traditional Medicine, Guangzhou University of Chinese Medicine, Guangzhou 510006, China
2.
South China Institute of Biomedicine, Guangzhou 510000, China
3.
College of Life Sciences, South China Agricultural University, Guangzhou 510642, China
4.
Zhuhai Hospital of Integrated Traditional Chinese and Western Medicine, Zhuhai, Guangdong 519000, China
5.
Yangchun Hospital of Traditional Chinese Medicine, Yangchun, Guangdong 529600, China
6.
Agricultural Experiment Farm of Yangchun City (Amomum villosum Testing Farm of Yangchun City), Yangchun, Guangdong 529600, China
Funds: This work was supported by grants from Forestry Science and Technology Innovation Project of Guangdong, China (2018KJCX033), Key-Area Research and Development Program of Guangdong Province (2020B020221002), Innovation Team Building Project of South Medicinal Industry in Modern Agricultural and Industrial Technology System, Guangdong China (2022KJ142; 2021KJ142; 2020KJ142; 2019KJ142), and National Key R&D Program of China (2017YFC1701102).
A series of cultivation experiments under different nutrient conditions were carried out on Amomum villosum Lour. plants through sand culture. The effects of nitrogen(N), phosphorus(P), potassium(K), calcium(Ca), boron(B), and zinc(Zn) concentrations on the growth and development of A. villosum were studied. Results showed that 15.0 mmol/L N, 1.0 mmol/L P, 6.0 mmol/L K, 0.5 mmol/L Ca, 8.0 × 10−5 mmol/L Zn, and 0.46 mmol/L B were sufficient for the growth and development of plants, and 10.0 mmol/L P, 8.0 × 10−3 mmol/L Zn, and 0.46 mmol/L B showed better performance. However, harmful symptoms appeared when the nutrient concentrations were increased to 50.0 mmol/L N, 6.0 × 102 mmol/L K, and 50.0 mmol/L Ca. Therefore, reasonable concentrations of mineral salts for fertilization should be used during the growth and development stages of A. villosum.
Amomum villosum Lour. is a perennial evergreen herb of the Zingiberaceae family[1]. The dried fruit of A. villosum is one of the “Four Famous Southern Chinese Medicines” (called Amomi Fructus), with an application history of more than 1300 years[2]. A. villosum is warm in nature and pungent in taste, and exerts various functions such as eliminating dampness, regulating stomach, warming spleen, antidiarrheal, regulating Qi, calming fetus[3]. The plant is primarily distributed in Guangdong, Guangxi, Yunnan, and Fujian provinces of China. Among these areas, Yangchun in Guangdong is considered the original production area of A. villosum, thus producing the best medicinal effects[4]. Market demand of the A. villosum industry is estimated to be more than 1.4 billion USD, however, unit yield of A. villosum is low and can not meet current demand[5]. The main reasons for this include low natural fruit setting rate, high fruit dropping rate, and artificial pollination difficulties[4, 6-8]. Various ecological factors, such as air humidity, can also influence A. villosum yield[9]. Furthermore, poor scientific management during the planting process and severe damage to the planting environment have also resulted in low reproduction and yield[9].
Successful reproductive growth of plants is based on good vegetative growth, especially during the seedling stage. Mineral salts play an important role in plant growth and development. There are 14 essential elements in soil, including six macroelements and eight microelements[10]. Nitrogen (N), phosphorus (P), and potassium (K) are the three main elements of plant nutrition and are essential for plant growth and development[11]. Calcium (Ca) maintains the structural stability of the membrane[10] and acts as a signaling factor to influence plant growth and development[12]. Zinc (Zn) is a component of many enzymes in plants, especially those involved in metabolic activities[13]. Boron (B) is an essential element for the formation and development of all nutritional and reproductive organs in plants[14]. To date, however, few studies have explored the effects of mineral salts on A. villosum growth and development. Notably, Zhang et al[15]. studied the effects of mineral salt deficiencies on growth and development of A. villosum seedlings over three months and found that 12 mineral salts exerted different effects on seedling growth, especially N, P, K, and iron (Fe). However, no research has reported on the patterns of demand for different mineral salts after A. villosum growth or how to reasonably apply these mineral salts. Currently, division propagation is commonly used in A. villosum production. With the growth and development of A. villosum ramet seedlings, the demand for various mineral salts, such as N, P, K, Ca, B, and Zn, will increase and also vary.
The aim of this study was to investigate the effects of abundance and deficiency of six mineral salts, including N, P, K, Ca, B, and Zn, on the growth and development of A. villosum plants and to explore the demand patterns and rational application of each mineral salt for seedling growth. This study should provide a theoretical basis for formulating a reasonable fertilization program for nutritional growth of A. villosum.
1.
Materials and Methods
1.1
Materials and their treatments
Amomum villosum cv. ‘Yuanguo’ seedlings were collected from Jilongkeng, Yangchun City, Guangdong Province, China. Plants showing vigorous growth (ca. 25 cm erect stems in height and ca. 20 cm young stolon buds in length) were selected as the initial culture materials. Whole plants were washed with tap water to remove soil from the roots, then rinsed twice with distilled water and planted immediately.
1.2
Experimental methods
1.2.1
Cultivation methods
The A. villosum seedlings were planted in blue plastic boxes (L∶W∶H = 70 cm∶48 cm∶35 cm) using a sand culture[16]. Three types of culture solutions were designed for cultivation: (1) complete nutrient solution; (2) amount of element 1/10 that of complete nutrient solution (defined as deficient nutrient solution, deficient treatment); (3) amount of element 10 times that of complete nutrient solution (defined as abundant nutrient solution, abundant treatment). There were 13 treatments, including complete nutrient solution (CK), 1/10 N (- N), 10 × N ( + N), 1/10 P (- P), 10 × P ( + P), 1/10 K (- K), 10 × K ( + K), 1/10 Ca (- Ca), 10 × Ca ( + Ca), 1/10 B (- B), 10 × B ( + B), 1/10 Zn (- Zn), and 10 × Zn ( + Zn) (Table 1). All plants were cultured in a greenhouse at Shi-Zhen Mountain on the campus of Guangzhou University of Chinese Medicine.
Table
1.
Ingredients of nutrient solutions used for treatments
Stock solution
Treatment
CK / mL
- N / mL
+ N / mL
- P / mL
+ P / mL
- K / mL
+ K / mL
- Ca / mL
+ Ca / mL
- B / mL
+ B / mL
- Zn / mL
+ Zn / mL
Ca(NO3)2 (0.5 mol/L)
10
10
10
10
10
10
10
10
10
KNO3 (0.5 mol/L)
10
10
10
10
10
10
10
10
10
MgSO4·7H2O (0.2 mol/L)
10
10
10
10
10
10
10
10
10
10
10
10
10
KH2PO4 (0.1 mol/L)
10
10
10
10
10
10
10
10
10
NaH2PO4 (0.1 mol/L)
1
100
10
10
Fe-EDTA (2 × 10−2 mol/L)
1
1
1
1
1
1
1
1
1
1
1
1
1
NaNO3 (0.5 mol/L)
1
100
10
10
10
10
CaCl2 (0.5 mol/L)
10
10
1
100
KCl (0.5 mol/L)
10
10
2
2
1.2
120
H3BO3 (4.6 × 10−2 mol/L)
1
1
1
1
1
1
1
1
1
0.1
10
1
1
CuSO4·5H2O (3 × 10−4 mol/L)
1
1
1
1
1
1
1
1
1
1
1
1
1
MnSO4 (6 × 10−3 mol/L)
1
1
1
1
1
1
1
1
1
1
1
1
1
Na2MoO4 (6 × 10−4 mol/L)
1
1
1
1
1
1
1
1
1
1
1
1
1
ZnSO4·7H2O (8 × 10−4 mol/L)
1
1
1
1
1
1
1
1
1
1
1
0.1
10
Note: The pH of the nutrient solution was adjusted to ~ 6.0 before use.
A river sand-vermiculite mixture (V∶V = 2∶1) was used as the cultivation substrate. A mixture of treated river sand (2/3 volume) and treated vermiculite (1/3 volume) was poured into a plastic box to a height of 20 cm and pressed firmly.
1.2.2.1
Pre-treatment of river sand
Small debris was removed using a stainless-steel sieve (5 mm aperture) and large debris was removed manually. The river sand was rinsed with water and soaked in 15%–18% hydrochloric acid and 1% oxalic acid for 3 d. After this, the sand was rinsed with water until neutral pH, then rinsed with distilled water three times and dried in the sun.
1.2.2.2
Pre-treatment of vermiculite
Vermiculite was soaked in water for 12 h, then rinsed and filtered using a nylon sieve (0.0075 mm aperture) until the effluent was clear and transparent. The vermiculite was then rinsed with distilled water three times after removing fine debris and dried in the sun.
1.2.3
Preparation of nutrient solution
The nutrient solution was prepared according to Hoagland’s formula for macroelements and Arnon’s formula for microelements. The chemical reagents used were of analytical grade. The macroelements were prepared with deionized water at 100 times the stock solution and stored separately; microelements were prepared with deionized water at 1000 times the stock solution and stored separately. Working nutrient solutions for the different treatments were prepared according to Table 1, and the total volume of each treatment solution was 1 L.
1.2.4
Planting and management
Before planting, the substrate was watered with 25 L of working nutrient solution to make the substrate slightly moist. The A. villosum plants were planted in the substrate, leaving the stolons exposed to the surface. Two plants were planted in each box and each treatment was repeated three times. The plants were watered with deionized water overnight for two weeks after planting, then watered once every two weeks at a dose of ~ 5 L of nutrient solution each time. After planting, the shed conditions were controlled at a temperature of 22℃–30℃, relative air humidity of 75%–90%, shade of 60%–80%, and soil moisture of 40%–46%.
1.2.5
Data measurement
The effects of different treatments on the growth and development of original A. villosum plants and new plants were determined. The physiological indices of the plants were measured, observed, and recorded every 10 d. Original plants were all plant parts at the time of collection, including original erect stems, leaves, stolons, and buds. New plants were plants differentiated from the stolons of the original plants, including newly differentiated erect stems, leaves, stolons, and buds. According to the A. villosum stolon grading method of He et al.[17], stolons and erect stems were divided into grades (0, 1, 2, 3, etc.). The youngest erect stem in a network was grade 0, and age increased with the number of grades.
1.2.5.1
Indicators of erect stem growth
Stem thickness: Diameter at 1/3 plant height was measured with vernier calipers. Plant height: Length of erect stems from the basal part of the stem to the base of the ligule of the uppermost leaf was measured with a soft tape measure.
1.2.5.2
Indicators of stolon growth
Stem thickness: Diameter of the stolon at the seventh node site (from erect stem at the corresponding grade) was measured with vernier calipers. Stem length: Distance between the basal parts of two adjacent erect stems was measured with a soft measuring tape. When no erect stems at the lower grade were found, the distance from the basal part of the stem to the bud tip was measured. Stem length was the sum of the length of the stolon at each grade. Number of buds: Number of young red buds longer than 1 cm on the stolon at each grade was counted. Number of nodes: Number of nodes on the stolon between two grades of erect stems was counted. Bud growth: Number of buds longer than 0.5 cm after planting was counted every 10 d. Bud growth was the difference between each count and number of buds on day 0 after planting. Node growth: Total number of stolon nodes at each grade after planting was counted every 10 d. Node growth was the difference between each count and number of nodes on day 0 after planting.
1.2.5.3
Indicators of leaf growth
Leaf length: Length of the middle leaf of plants with more than five leaves was measured. Middle leaf was defined as the n/2nd leaf (from the base of the plant) when the number of leaves of the plant was n and n was an even number, or (n + 1)/2nd leaf (from the base of the plant) when n was an odd number. The length of the middle leaf was measured with a plastic ruler and repeated three times. Number of leaves: Number of fully expanded leaves on an erect stem was counted. Chlorophyll content: Chlorophyll content in the middle leaves on erect stems was determined with a rapid chlorophyll meter (Chlorophyll Meter SPAD-502, Konnica Minolta Sensing, Inc.) and repeated three times. Leaf growth: Number of leaves of erect stems at all grades was counted every 10 days after planting. Leaf growth was the difference between each count and number of leaves on day 0 after planting.
1.2.5.4
Observation of plant characteristics
Plant softening and wilting, color changes on different parts, and recoiled leaf length and proportion to total leaf length were observed and recorded.
1.2.5.5
Observation of pest and disease symptoms
Symptoms of pests and diseases on the plants were observed, recorded, and identified.
1.3
Statistics
Data for each growth index of the A. villosum plants were processed and plotted using Excel 2019 software. Statistical analysis was performed by one-way analysis of variance (ANOVA) using SPSS v20.0 software. Data are summarized as mean ± SEM.
2.
Results
2.1
Effects of different treatments on growth of original A. villosum plants
2.1.1
Effects of different treatments on growth of erect stems of original A. villosum plants
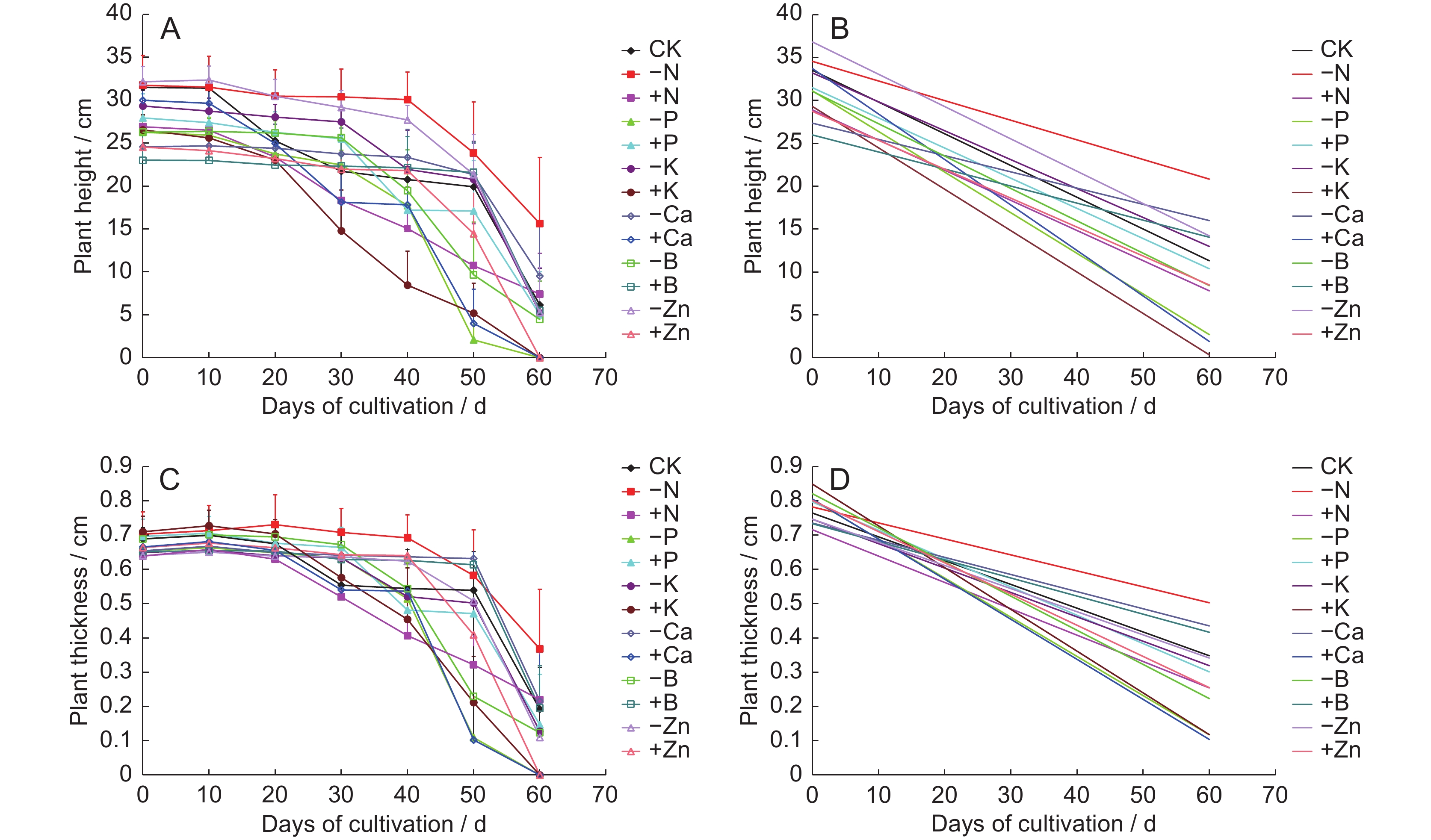
The growth of erect stems of original plants was assessed in response to six mineral salts under abundant and deficient conditions in nutrient solution. The plant height of each treatment group showed a negative growth trend (Fig.1: A), whereby erect stems gradually senesced and withered from the physiological top downwards. Plants in the - P, + K, + Ca, and + Zn treatment groups completely withered and died by day 60. To simplify the results, we set a linear trend line for each treatment (Fig.1: B) with the linear function y = kx + b (y: plant height, x: number of days in cultivation, b: constant, k: slope), with k representing the rate of erect stem senescence and wilting. The absolute value of k for the 13 treatments was in the order: + Ca > + K > - P > (- B, - Zn, CK) > ( + P, + N) > ( + Zn, - K) > - N > + B > - Ca. Therefore, under abundant Ca and K treatment and deficient P treatment, erect stems senesced faster than under CK treatment. In contrast, the rate of senescence was slower under abundant Zn treatment than under CK treatment until day 40 of cultivation. However, erect stems showed sudden accelerated senescence after 40 d of cultivation and wilted and died by day 60.
Figure
1.
Effects (A, C) and effect trends (B, D) of different treatments on height and diameter of erect stems of original plants
Erect stem thickness showed a negative trend in all treatment groups (Fig.1: C), suggesting that erect stems had lost water and withered. Plants in the - P, + K, + Ca, and + Zn treatment groups were dead by day 60, and thus stem thickness could not be measured. As with the simplified treatments above, slope k of the linear function represented the shrinkage rate of erect stems (Fig.1: D). The absolute value of k for the 13 treatments was in the order: + K > ( + Ca, - P) > (- B, + N) > + Zn > + P > - K > (CK, - Zn) > ( + B, - Ca) > - N. Similar to the changes in erect stem height, abundant Ca and K treatment and deficient P treatment accelerated shrinkage of erect stem thickness. In contrast, abundant B treatment and deficient Ca and N treatment delayed this shrinkage.
All original plants senesced and died after transplantation, but the rate of senescence differed under different mineral salt treatments (Fig.1). Abundant Ca and K treatment and deficient P treatment accelerated senescence and death, while abundant B treatment and deficient Ca and N treatment delayed the senescence process. Abundant Zn treatment delayed senescence for the first 40 d after transplantation but accelerated senescence after 40 d.
2.1.2
Effects of different treatments on germination of new buds of original A. villosum plants
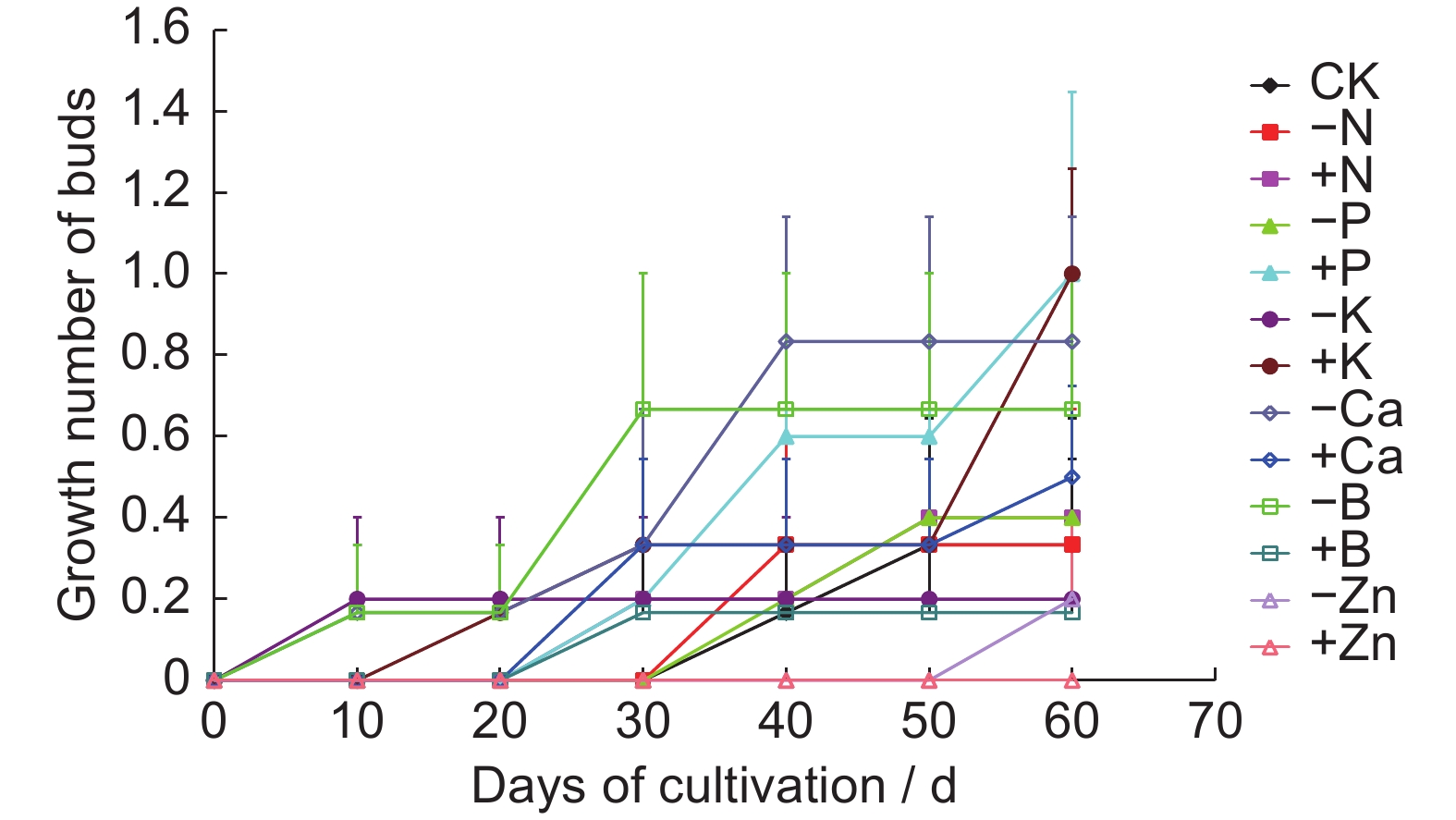
The average number of germinations under high P and K treatment reached one after 60 d, with significantly higher abundance of new buds compared to other treatments and germination times of 30 and 20 d after transplantation, respectively (Fig.2). Deficient Ca treatment produced the next most abundant buds, with a germination time of 10 d after transplantation. On day 60, no new buds germinated under abundant Zn treatment and only one new bud was found under deficient Zn treatment, indicating that the addition of Zn inhibited new bud germination. Similarly, abundant B treatment also inhibited new bud germination.
Figure
2.
Effects of different treatments on germination of new buds of original plants
2.1.3
Effects of different treatments on growth of stolons of original A. villosum plants
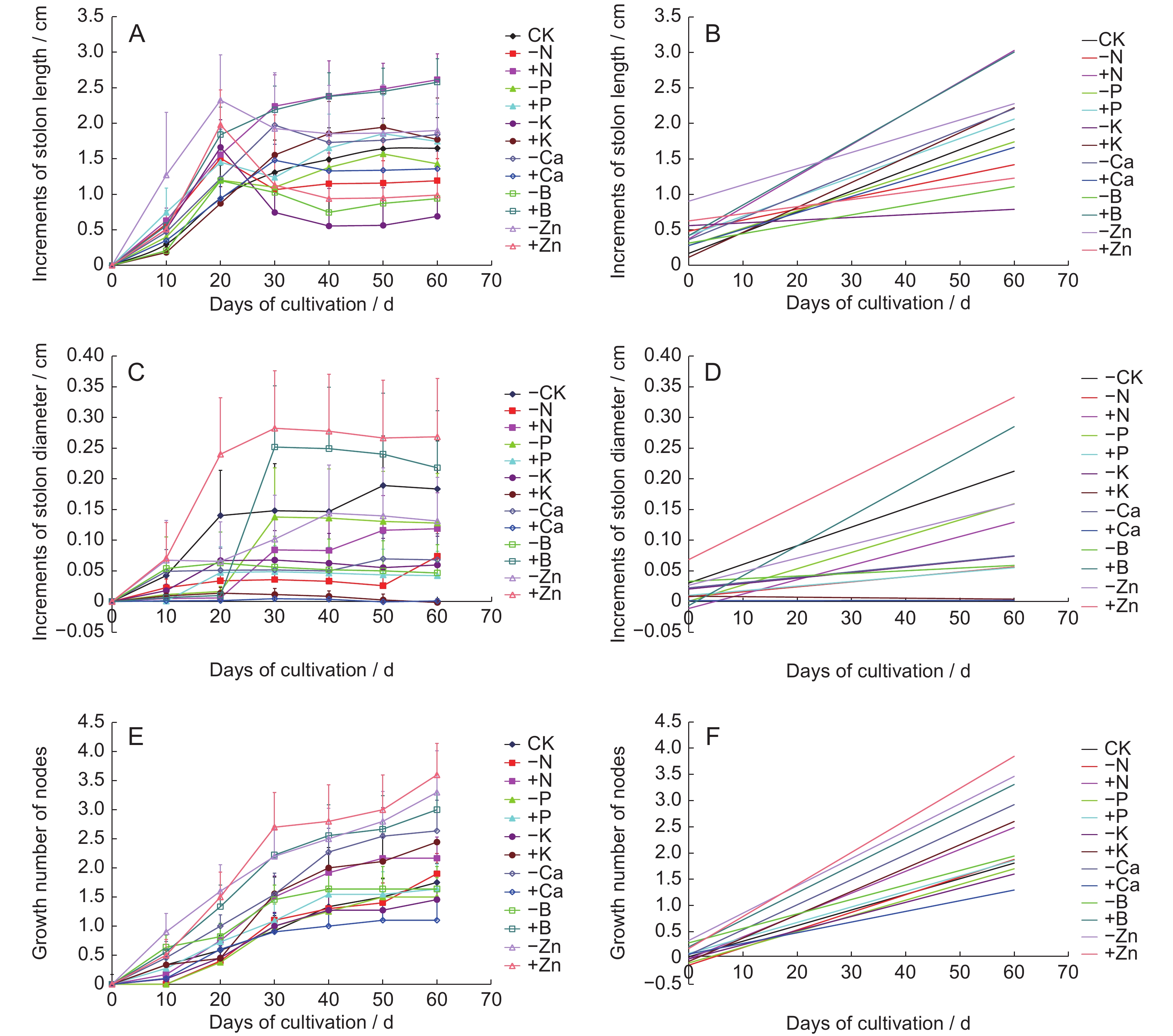
Each treatment group showed a certain promoting effect on the elongation of stolons and differentiation of nodes of the original A. villosum plants. Treatments either promoted or inhibited stem thickness of stolons; however, there was no significant difference among treatment groups (Fig.3).
Figure
3.
Effects (A, C, E) and effect trends (B, D, E) of different treatments on growth of stolons of original plants
The + K treatment promoted stolon length growth in the original plants, with a maximum growth rate of 43.07% (Fig.3: A) and more pronounced effects in the earlier and middle stages. In addition, - N, - K, and - Ca promoted stolon diameter growth, with - N inducing the highest growth rate of 3.87% (Fig.3: C) but only in the earlier stage of cultivation. The + B treatment exerted a strong effect on stolon thickness growth within a certain period, with a peak on day 30 and the highest rate (0.3025%) among all treatments (Fig.3: C). Stolon thickness decreased with the accumulation of + B in the soil, resulting in a negative growth rate of −4.67% (Fig.3: C). The effects of + Zn, - Zn, and + B on stolon nodal growth were significant, with + Zn showing the most obvious effects (3.60 nodal growth) (Fig.3: E).
2.2
Effects of different treatments on growth and development of new A. villosum plants
2.2.1
Effects of different treatments on growth and development of erect stems of new plants
Observations of growth and development on day 60 showed that the new A. villosum plants in the - N, - P, + P, - K, - Ca, - B, + B, - Zn, + Zn, and CK treatment groups grew relatively healthy, while plants in the + N and + K treatment groups were stunted and plants in the + Ca treatment group were decayed (Fig.4).
Figure
4.
Effects of different treatments on growth of erect stems of new plants
A1: - N treatment; A2: + N treatment; B1: - P treatment; B2: + P treatment; C1: - K treatment; C2: + K treatment; D1: - Ca treatment; D2: + Ca treatment; E1: - B treatment; E2: + B treatment; F1: - Zn treatment; F2: + Zn treatment; G: CK treatment. Same below.
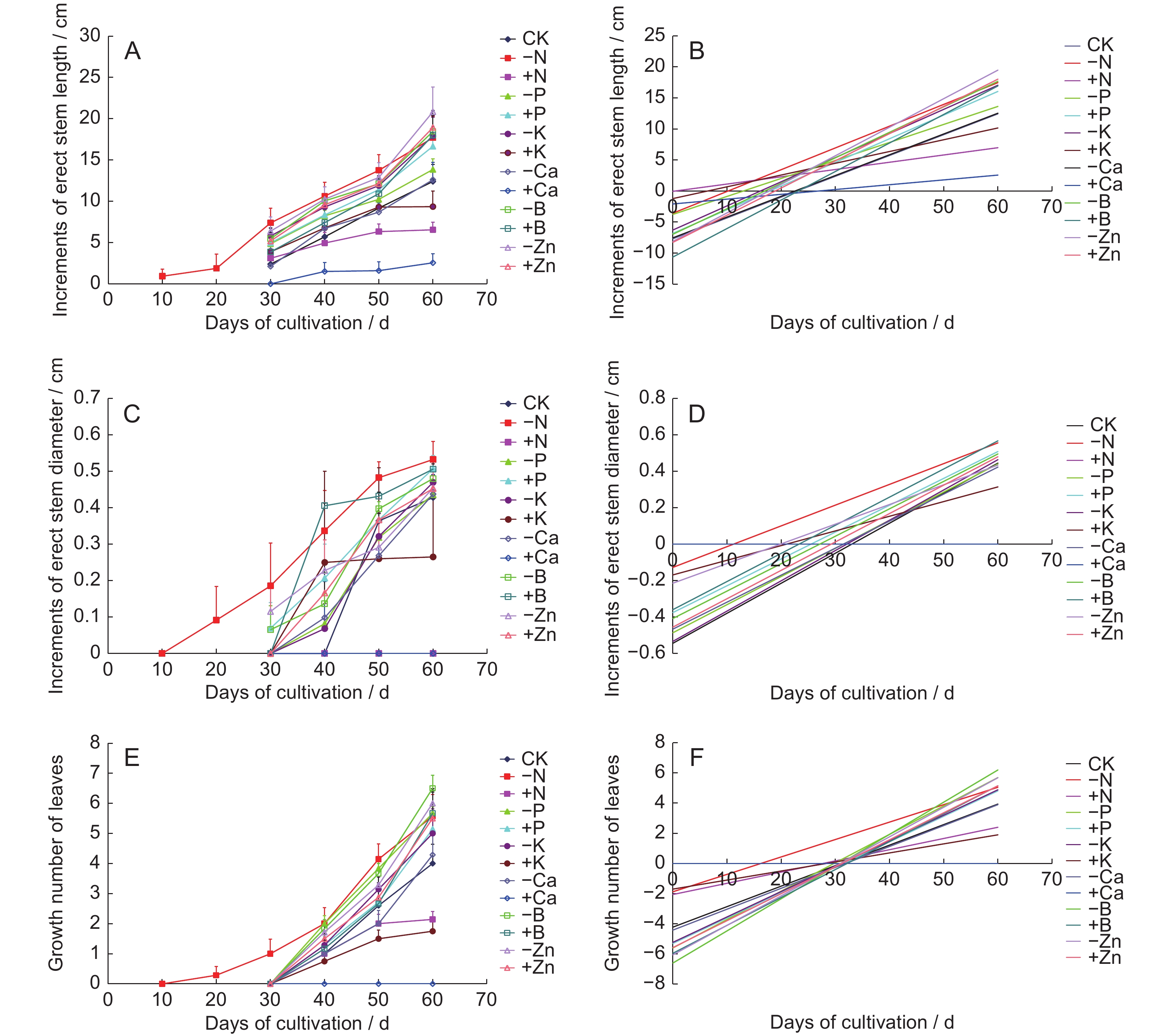
Different treatments had different effects on plant height, stem thickness, and differentiation time after planting. Among the groups, - Zn treatment showed the highest new plant height growth (20.84 cm) and better treatment effects, with growth occurring in the later stage of cultivation, increasing by 7.91 cm from day 50 to 60 (Fig.5: A, B). For stem thickness, the - N group showed a continuous growth trend, while the growth in all other treatment groups occurred in the middle and later stages (Fig.5: C, D). Stem thickness growth did not vary significantly in each treatment group, with a maximum value of 0.53 cm (Fig.5: C). The + Ca treatment was toxic to A. villosum, causing rapid decay of the original plants and inhibiting the differentiation and growth of new plants, so statistical comparisons could not be made.
Figure
5.
Effects (A, C, E) and effect trends (B, D, F) of different treatments on growth of erect stems and leaf blades of new plants
The - N treatment accelerated differentiation of new erect stem organs in the early stage. However, - N treatment had a negative effect on organ development in the late stages, with plants showing excessive growth in the earlier stage but slow growth in the later stage, resulting in thin seedlings. The - Zn treatment was more effective for the establishment of new plants, with a faster growth rate in the later stage. Although induction time in the + B treatment group was long, it promoted the growth of plant height and stem thickness in the later stage.
2.2.2
Effects of different treatments on leaf growth and development of new plants
The growth and development of new plant leaves on day 60 was observed (Fig.6). Among the effects of each treatment group on leaf growth and development of new plants, the - N treatment showed initial leaf growth on day 10. However, leaf growth under all other treatments appeared after day 30 (Fig.5: E, F).
Figure
6.
Effects of different treatments on growth of leaf blades of new plants
There were some differences among the treatments in indicators of new plant leaf growth and development. The - B treatment produced the greatest number of leaves (7.17) (Table 2). The + B treatment produced the longest leaves (13.65 cm) and highest chlorophyll content (46.93) (Table 2). The - Ca treatment had the fastest effect on leaf formation and growth promotion. Element deficiency induced rapid formation of new organs but was detrimental to later growth and development. Although + B required more time to induce leaf differentiation, this treatment group contained well-developed long leaves with high chlorophyll content, which had a positive effect on photosynthesis.
Table
2.
Effects of different treatments on growth of leaf blades of new plants
Treatment
Number of new plants
Number of leaf blades
Leaf length / cm
Chlorophyll content (SPDA value)
Leaf differentiation time / d
CK
5
4.20 ± 0.73cd
9.30 ± 0.36a
37.48 ± 2.82bcd
8.00 ± 2.00ab
- N
7
5.57 ± 0.72abc
11.48 ± 1.50a
31.68 ± 1.56d
7.14 ± 1.84ab
+ N
7
2.29 ± 0.18d
−
−
4.29 ± 2.02ab
- P
6
5.83 ± 0.17abc
9.67 ± 0.83a
30.96 ± 3.54d
8.33 ± 1.67a
+ P
7
5.86 ± 1.01abc
12.74 ± 0.56a
44.87 ± 3.22ab
1.43 ± 1.43b
- K
7
5.86 ± 0.86abc
11.24 ± 0.48a
44.27 ± 1.92ab
1.43 ± 1.43b
+ K
4
2.25 ± 0.48d
−
−
2.50 ± 2.50ab
- Ca
7
4.71 ± 0.92bc
10.03 ± 1.14a
34.92 ± 5.11cd
0.00 ± 0.00ab
+ Ca
1
−
−
−
−
- B
6
7.17 ± 0.60a
10.80 ± 0.69a
42.09 ± 2.74abc
4.00 ± 2.45ab
+ B
6
6.17 ± 0.91abc
13.65 ± 0.89a
46.93 ± 0.64a
8.00 ± 2.00ab
- Zn
7
6.71 ± 0.57ab
11.44 ± 1.20a
40.09 ± 1.76abc
2.86 ± 1.84ab
+ Zn
8
6.00 ± 0.98abc
12.00 ± 0.14a
43.71 ± 1.12ab
4.29 ± 2.02a
Notes: All data are mean ± SEM. Single identical letter in the same column indicates no significant difference at 0.05 level. Same below.
The - Ca treatment was the most effective in promoting bud differentiation, inducing differentiation within the shortest amount of time (20 d). The - B treatment had the best effect on stolon stem length growth (maximum of 5.80 cm) and was significantly different from the CK treatment but not from the other groups. The + P treatment had a more significant effect on the number of buds, stolon length, and differentiation time of buds in new plants (Table 3).
Table
3.
Effects of different treatments on differentiation in new plants
As a traditional medicinal herb in China, A. villosum is one of the first eight medicinal herbs protected by legislation in Guangdong Province. Appropriate fertilization can ensure vigorous plant grow and improved fruiting and fruit setting rates, thereby increasing A. villosum yield[9]. Therefore, investigating the effects of mineral salt abundance and deficiency on A. villosum growth and development is crucial to formulate a reasonable fertilization plan to ensure good growth and yield. This study found that different concentrations of six mineral salts had different effects on the growth and development of both original and new plants.
3.1
Effects of macroelements on growth and development of A. villosum plants
Macroelements (C, H, O, N, P, S, K, Ca, and Mg) are essential for plants[10]. In this study, we investigated the effects of four of these macroelements (N, P, K, and Ca) on the growth and development of A. villosum seedlings.
Our results showed that -N treatment inhibited the decay of erect stems of the original plants. Furthermore, this treatment promoted stolon thickening in the original plants as well as the formation and pre-growth of erect stems of new plants. However, the effects were not significantly different from those of other treatment groups. In addition, - N treatment caused the new plants to grow slowly at the later stage, and plants showed symptoms of element deficiency with obvious chlorosis, similar to the results of Zhang et al.[15] We also found that - K treatment inhibited stolon growth (length and diameter) of the original plants but had no significant effect on the growth and development of new plants. This is consistent with Chen et al.[18], where low K treatment was shown to inhibit plant roots. Ca salt helps maintain normal physiological activities of leaves and promote leaf growth[19]. In this study, - Ca treatment effectively induced the formation of new leaves and buds in A. villosum plants but prolonged the establishment time of new plants. During later growth and development, - Ca treatment showed better results than the + Ca and CK treatment groups.
The + N, + K, and + Ca treatments all showed different degrees of toxicity during A. villosum growth and development, with poor plant development and even decay. For most plants, excessive N fertilization can promote the synthesis of auxin and cytokinin, which can cause plant overgrowth and delayed maturity[20]. In the present study, however, the plants showed dwarfing, chlorosis, and withering. This has not been reported in other crops and the reasons need to be further explored. High K treatment hindered plant growth, probably because higher K levels can disturb ionic balance in the plant, thereby impacting chlorophyll synthesis, causing a decrease in photosynthesis and dysregulation of sugar metabolism[21]. Excessive Ca uptake can also cause stomatal closure of leaves, which can inhibit plant growth[22].
The effect of P treatment on A. villosum plants was not obvious. However, it still promoted erect stem length and diameter in new plants, as well as the formation and later development of new leaves and induction of subordinate stems in new plants. Furthermore, + P treatment promoted new bud growth better than the - P and CK treatments.
3.2
Effects of microelements on growth and development of A. villosum plants
Plant demand for microelements (Cl, Fe, B, Mn, Zn, Cu, Ni, and Mo) is extremely low, and slightly higher doses can have toxic effects on plants[10]. In this study, we investigated the effects of two of these microelements (B and Zn) on A. villosum growth and development.
The results showed that - B treatment significantly promoted leaf number and stolon length in the new plants. The + B treatment significantly increased chlorophyll content in the leaves, consistent with the findings of Li et al.[20] on jasmine. Zn not only activates enzymes but is also a component of many enzymes and chlorophyll, resulting in an increase in photosynthesis under increasing concentrations[22]. Here, - Zn treatment significantly promoted erect stem growth (length and diameter) and earlier stage differentiation in the new plants. The + Zn treatment also promoted the growth of nodes, leaves, and chlorophyll content of the original plants, provided nutrients for later growth, and significantly promoted the growth of subordinate stems.
4.
Conclusions
Considering the changes in the plant growth indices from days 0 to 60, we recommend the following optimal amounts of each salt for fertilization of A. villosum plants: 15.0 mmol/L N, 10.0 mmol/L P, 6 mmol/L K, 5.0 × 10−1 mmol/L Ca, 8.0 × 10−3 mmol/L Zn, and 4.6 × 10−1 mmol/L B.
As actual soil environments and rainfall will differ from the cultivation environment in this study, the addition of salts may not achieve the same effects as in this study. Thus, the aforementioned optimal doses form the basis of fertilization and can be increased appropriately, although at levels below 10 times the dosage to avoid waste and physiological toxicity to plants, and applied once every 1–2 months.
Lü BD, Hu JJ, Tang LY, Su J, Xu J, He GZ. The study on the fruit dropping law and physiological mechanism of Amomum villosum[J]. Plant Physiology Communications, 2021, 57(2): 429-438.
Tang LY, He GZ, Su J, Xu HH. The strategy to promote the development of industry of genuine medicinal material of Amomum villosum[J]. Chinese Agricultural Science Bulletin, 2012, 28(8): 94-99. doi: 10.3969/j.issn.1000-6850.2012.08.021
[5]
Yang R,Wang J,Gao W,Jiang Y,Su J,et al. Research on the reproductive biological characteristics of Amomum villosum Lour. and Amomum longiligulare T. L. Wu.[J]. PLoS One,2021,16 (8):e0250335. doi: 10.1371/journal.pone.0250335
He GZ, Gao W, Su J, Li JK, Tang LY. Floweret configuration characters of the medicinal plant Amomun villosum[J]. Bulletin of Botany, 2014, 49(3): 313-321. doi: 10.3724/SP.J.1259.2014.00313
[9]
Xu J, Li MX, Su J, Tang LY, Jiang Y, et al. Study on ecological stereoscopic cultivation mode of Amomum villosum-Dimocarpus longan[J]. China Journal of Chinese Materia Medica, 2018, 43(2): 288-298.
[10]
王小菁. 植物生理学[M]. 北京: 高等教育出版社, 2019: 33−36.
[11]
陆欣. 土壤肥料学[M]. 北京: 中国农业大学出版社, 2002: 167.
[12]
Raina M,Kumar A,Yadav N,Kumari S,Yusuf MA,et al. StCaM2,a calcium binding protein,alleviates negative effects of salinity and drought stress in tobacco[J]. Plant Mol Biol,2021,106 (1-2):85−108. doi: 10.1007/s11103-021-01131-1
[13]
Hassanein YZ,Abdel-Rahman SSA,Soliman WS,Salaheldin S. Growth,yield,and quality of roselle (Hibiscus sabdariffa L.) plants as affected by nano zinc and bio-stimulant treatments[J]. Hortic Environ Biotechnol,2021,62 (6):879−890. doi: 10.1007/s13580-021-00371-w
Zhang LX, Peng JM, Gao WW, Ma J, He YQ. Effect of nutritional deficiency on the growth of Amomum villosum seedling[J]. Journal of Yunnan Agricultural University, 2011, 26(5): 700-705.
[16]
毛达如. 植物营养研究方法[M]. 2版. 北京: 中国农业大学出版社, 2005: 25−29.
[17]
He GZ, Gao W, Su J, Li JK. Differentiation rule of Amomum villosum Lour. floral bud[J]. International Medicine and Health Guidance News, 2012, 18(15): 2142-2144.
[18]
Chen G, Gao ZY, Xu GH. Adaption of plants to potassium deficiency and strategies to improve potassium use efficiency[J]. Bulletin of Botany, 2017, 52(1): 89-101. doi: 10.11983/CBB16231
[19]
Dong WC, Liu XB, Li HM, Zhao SM, Bao JM, et al. Effects of calcium supply with varying levels on the growth and development of woody ornamental plants[J]. Chinese Agricultural Science Bulletin, 2022(8): 42-50.
[20]
Li CN, Huang ZW, Li XM, Lu JS, Bu ZY, et al. Effects of combined fertilization of zinc, boron and molybdenum on flowering and leaf nutrients of jasmine (Jasminum sambac)[J]. Soil and Fertilizer Sciences in China, 2022(5): 1-7.
[21]
Gao WQ, Liu CY, Wu QS. Effects of potassium stress on growth, phytohormone and signal substance levels in roots of trifoliate orange[J]. Jiangsu Agricultural Sciences, 2022(8): 42-50.

 DownLoad:
DownLoad: